Why David Pearce's "Evidence for Design" fails to disprove evolution - 3
One of the most readily-refuted
creationist claims is the absence of transitional fossils. Transitional forms
abound in the fossil record, and provide compelling evidence for large-scale
evolutionary change. [1] Therefore, when Pearce declares that:
There are also enormous problems at the top end of the diagram. If life progressed steadily from simple forms to complex ones, where are the intermediate stages now? Why cannot their remains be found in the fossil rocks? [2]
not only is he showing that he
fails to grasp the fact that evolution is not a linear progression from microbe
to man, but a bushy branch as one would expect from a process of descent with
modification, he is positively telegraphing his lack of understanding of palaeontology to his
informed readers.
His insistence that evolution
should have produced intermediate forms reflects a fundamental misunderstanding
about evolutionary biology. As mentioned, evolution does not proceed from
simple to complex along a line, but is a branching process. Therefore, we do not
expect to see something halfway between a modern reptile and a modern mammal.
Living reptiles and living mammals share a common ancestor that lived around
325 million years ago [3] so it is
a mistake to think that palaeontologists specialising in mammalian evolution
are looking for a ‘missing link’ that looks like an intermediate between modern
mammals and modern reptiles. Louise Mead notes that:
Expectations of “intermediate” forms reflect neither Darwin’s original thoughts nor current thinking and practices in evolutionary biology and paleontology. Darwin (1859) was very clear on this. The ancestor of two living forms is unlikely to be found alive because it would have been outcompeted in most cases by newly adapting forms, and an extinct ancestor of two living forms would not be expected to look intermediate between them. Today, evolutionary biologists and paleontologists do not focus on finding “intermediates” but rather on reconstructing evolutionary relationships and history using shared derived characters or synapomorphies... Willi Hennig revolutionized systematics in the 1960 s with the introduction of cladistics, which ushered in a new method of phylogenetic analysis and a new approach to systematics. Instead of relying on a Linnaean system of classification, cladistics placed the focus on evolutionary history, specifically identifying features as ancestral (general) or derived (evolved after the lineage split from the ancestor). If a shared derived character or synapomorphy is found in two or more related organisms, it is inferred to have been present in their common ancestor, irrespective of whether or not there is a fossil record for that ancestor. Rather than trying to find the actual fossil corresponding to the “missing link” between lobe-fins and tetrapods, paleontologists instead look for fossils with characters or features important for an adaptive transition from life in an aquatic environment to life on land and that are shared as the result of common ancestry. [4]
If we look at the consensus cladogram
of tetrapodomorphs, we can see that palaeontologists do not think that tetrapod
evolution is a linear process from fish to tetrapod. Rather, we see fossils
with those features that reflect a transition from aquatic to terrestrial life,
such as limbs capable of moving on dry land, and a transition towards
breathing.
 |
Evo Edu Outreach (2009) 2:213–223
|
The transition from a aquatic
existence to a terrestrial existence is well documented as evidenced by a
splendid series of transitional forms, which I will document in some detail. No
apology is made for this – Pearce has claimed that there are no intermediates,
and the best way to show how utterly uninformed he is on the subject is to show
in detail why this claim is wrong.
Kenichthys: is a 395 million year
old tetrapodomorph fossil that shows evidence of how the nostrils evolved from
the paired fish configuration to the internal/external tetrapod configuration.[5] Most living fish have paired nostrils that do
not connect with the internal cavity. The anterior nostril allows water to flow
in, while the posterior nostril allows water to flow out. In tetrapods, there is
only one pair of external nostrils, with the choanae connecting the internal
nostrils to the nasal cavity. Embryological studies show that these ‘internal
nostrils’ of lungfish are homologous to the posterior nostrils of fish. This
lends support to the idea that the posterior nostrils moved and became the
choanae in tetrapods:

FIGURE 1. Nostril positions on the heads of sarcopterygian fishes.
a, reconstruction of Kenichthys campbelli head and cheek in lateral view. b, c, Youngolepis. d, e, Kenichthys. f, g, Eusthenopteron. b, d, f are lateral views; c, e, g are ventral views. In e, the (unknown) vomer is represented by its attachment area on the ethmoid. Not to scale. ch, choana; Dpl, dermopalatine; Enpt, entopterygoid; La, lacrimal; l.Ro, lateral rostral; Mx, maxilla; n.a, anterior nostril; n.p, posterior nostril; Pmx, premaxilla; Te, tectal; Vo, vomer. - Zhu M, Ahlberg PE "The origin of the internal nostril of tetrapods" Nature (2004) 432:94-97 doi:10.1038/nature02843

FIGURE 2. Photographs of Kenichthys campbelli specimens.
a, b, V10493.61, ethmosphenoid in ventral (a) and anterior (b) views. c, d, V10493.60, ethmosphenoid in ventral (c) and lateral (d) views. e, V10493.58, ethmosphenoid in antero-lateral view. f, V10493.1, holotype, ethmosphenoid in antero-lateral view. g-h, V10493.101, ethmosphenoid in anterior view. i, V10493.81, lacrimal in external view. j, V10493.102, ethmosphenoid in anterior view. k, V10493.77, maxilla in external view. l, V10493.76, maxilla in external view. The green arrow indicates the free ventral margin of the lateral rostral; the red arrow in i indicates the slight 'step' marking the boundary of the tectal and prefrontal sutures. Scale bar, 2 mm. Zhu M, Ahlberg PE "The origin of the internal nostril of tetrapods" Nature (2004) 432:94-97 doi:10.1038/nature02843
a, b, V10493.61, ethmosphenoid in ventral (a) and anterior (b) views. c, d, V10493.60, ethmosphenoid in ventral (c) and lateral (d) views. e, V10493.58, ethmosphenoid in antero-lateral view. f, V10493.1, holotype, ethmosphenoid in antero-lateral view. g-h, V10493.101, ethmosphenoid in anterior view. i, V10493.81, lacrimal in external view. j, V10493.102, ethmosphenoid in anterior view. k, V10493.77, maxilla in external view. l, V10493.76, maxilla in external view. The green arrow indicates the free ventral margin of the lateral rostral; the red arrow in i indicates the slight 'step' marking the boundary of the tectal and prefrontal sutures. Scale bar, 2 mm. Zhu M, Ahlberg PE "The origin of the internal nostril of tetrapods" Nature (2004) 432:94-97 doi:10.1038/nature02843
Kenicthys is clearly a tetrapodomorph, and it displays exactly the intermediate condition that would be predicted when assuming that the tetrapod choanae were posterior nostrils displaced into the palate, in the same way as in lungfishes. The posterior nostrils of Kenichthys are actually right at the margin of the upper jaw, interrupting the outer dental ‘arcade’ formed by the premaxillary and maxillary bones.
Kenichthys clearly provides the first factual basis for the theory that tetrapod choanae actually are the posterior nostrils, as are the internal nostrils of lungfish. [6]

FIGURE 3. Morphology of Kenichthys.
a–f, h, drawings of Kenichthys campbelli specimens. a, Restoration of ethmosphenoid in ventral view, based mainly on c and d. b, c, V10493.61, ethmosphenoid in anterior (b) and ventral (c) views. d, e, V10493.60, ethmosphenoid in ventral (d) and lateral (e) views. f, V10493.77, maxilla in external view. g, Maxilla of Medoevia (ref. 15), showing the diverging processes framing the choana. h, V10493.58, ethmosphenoid in antero-lateral view. Scale bar, 2 mm. - Zhu M, Ahlberg PE "The origin of the internal nostril of tetrapods" Nature (2004) 432:94-97 doi:10.1038/nature02843

FIGURE 1. The hole picture.
These depictions of the head and palate in some living and fossil lobe-finned fishes and tetrapods show the apertures of the nasal capsule — the anterior (a) and posterior (p) nostrils and the choanae (c). The tree indicates the relationships of the groups, and the orange arrows indicate the two occurrences of the migration of the posterior nostril into the palate. In Kenichthys, the fossil studied by Zhu and Ahlberg1, the posterior nostril forms a notch in the outer dental arcade, and is intermediate in position between that of an external nostril and that of choanae (p/c). The diagrams (1–3) on the right are of the left nasal cavity, in ventral view, in the three living forms. In lungfishes, the path of the maxillary nerve (mx) is medial to the posterior nostril; in tetrapods it is lateral to the choana. t, tear duct. - Janvier P "Wandering Nostrils" Nature (2004) 432:23-24 doi:10.1038/432023a
These depictions of the head and palate in some living and fossil lobe-finned fishes and tetrapods show the apertures of the nasal capsule — the anterior (a) and posterior (p) nostrils and the choanae (c). The tree indicates the relationships of the groups, and the orange arrows indicate the two occurrences of the migration of the posterior nostril into the palate. In Kenichthys, the fossil studied by Zhu and Ahlberg1, the posterior nostril forms a notch in the outer dental arcade, and is intermediate in position between that of an external nostril and that of choanae (p/c). The diagrams (1–3) on the right are of the left nasal cavity, in ventral view, in the three living forms. In lungfishes, the path of the maxillary nerve (mx) is medial to the posterior nostril; in tetrapods it is lateral to the choana. t, tear duct. - Janvier P "Wandering Nostrils" Nature (2004) 432:23-24 doi:10.1038/432023a
Osteolepis: 390 million year old
lobe-finned fish that has paired front and rear fins that has limb-like
arrangement of the bones in its lobe fins, internal nostrils and amphibian-like
skull and teeth. From an evolutionary point of view, its primitive middle ear
is tetrapod-like in its configuration. This is a fish which has tetrapod
features in its fins, skull and ears:

Osteolepis macrolepidotus fossil at the Museum für Naturkunde, Berlin. (Source: Wikipedia)
A specimen of Osteolepis macrolepidotus from the Middle Old Red Sandstone of Tynet Burn, Banffshire, in the Museum of the Zoological Department of University College, shows with perfect clearness a cast of the cavities left in the completely ossified otic capsule of both sides. The specimen is illustrated in the photograph.
The three semicircular canals, which automatically analyse any movement into three components referred to rectangular and rectilinear coordinates, form a theoretically perfect mechanism and have persisted without any serious modification through the vertebrates. [7]

Fig 1. Head of Osteolepis macrolepidotis, x 2 approx. (J Anat. 1925 July; 59(Pt 4): 385–386.)
Olf.C. olfactory capsule; A.C.S.A. ampulla of anterior vertical semicircular canal;
A.C.S.H. ampulla of horizontal canal; C.S.P. posterior vertical semicircular canal;
Hy. Md. Hyomandibular
Note that this reference dates to 1925. Given its antiquity, Pearce has no excuse for failing to inform his readers of the existence of this fossil. If he had properly researched his subject and consulted experts in the area, he would have been well aware of this transitional fossil. Once again, it shows that he is not in a position to make any authoritative comments on this subject.
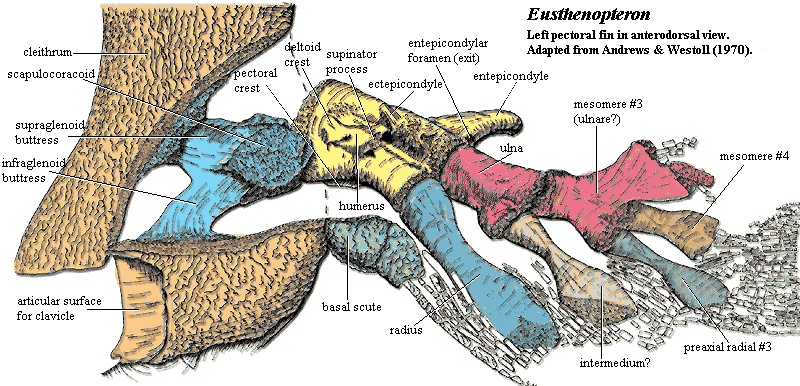
Eusthenopteron: 385 million year old lobe finned fish which has many transitional features indicating that it was well on the way to becoming a tetrapod.

Source: Wikipedia
- It has paired fins with distinct ulna, radius and humerus in the front fins, and fibula, tibia and femur in the rear fins. These are the arm and leg bones in tetrapods like us.
- It has internal nostrils
- It has teeth with infolded enamel such as seen in tetrapods

Source: Wikipedia
Clack comments on Eusthenopteron, noting that while it has significant tetrapod-like features, it also has fish-like characteristics:
A feature shared by most fishes is the bone called the hyomandibula that supports the palate and controls movements of the gill apparatus. It is a long bone that pivots on the walls of the bone housing the brain and ear capsules, the braincase, and it has attachment points to the gill bars and the opercular series, the plate-like bones that cover and protect the gills. They are all part of the pumping mechanism used in breathing and feeding. Eusthenopteron was entirely typical of early fish in this respect. [8]


Source
The presence of fin rays differentiates limbs from fins, so Eusthenopteron is viewed as a fish. However, its fins have strong tetrapod-like features – forelimb bones homologous to the humerus, ulna and radius. It is a fish with limbs but no hands or feet. Creationists who talk about the creation of fixed ‘kinds’ in which limited evolution occurs do not have the luxury of resorting to this tactic, as objectively, this fossil has features that are fish-like, and features that are tetrapod-like. The presence of fin rays is one of the diagnostic criteria for defining a taxon as fish, so special creationists are fond of dismissing such fossils as fish with mosaic characteristics. However, that is exactly what palaeontologists regard this lobe-finned fish. A lobe-finned fish with a mosaic of tetrapod and fish characteristics is a fossil with transitional features.
The presence of fin rays differentiates limbs from fins, so Eusthenopteron is viewed as a fish. However, its fins have strong tetrapod-like features – forelimb bones homologous to the humerus, ulna and radius. It is a fish with limbs but no hands or feet. Creationists who talk about the creation of fixed ‘kinds’ in which limited evolution occurs do not have the luxury of resorting to this tactic, as objectively, this fossil has features that are fish-like, and features that are tetrapod-like. The presence of fin rays is one of the diagnostic criteria for defining a taxon as fish, so special creationists are fond of dismissing such fossils as fish with mosaic characteristics. However, that is exactly what palaeontologists regard this lobe-finned fish. A lobe-finned fish with a mosaic of tetrapod and fish characteristics is a fossil with transitional features.
Gogonasus: this is a lobe finned fish that has a tetrapod-like middle ear and bones in its pectoral fins that were tetrapod-like. Its skeletal anatomy [9] shows that it was more tetrapod-like than Eusthenopteron.

FIGURE 1. Cranial features of Gogonasus andrewsae, NMV P221807.
a, Skull in lateral view. b, c, skull in dorsal view. d, e, X-ray micro-tomograms showing the spiracular region in lateral (d) and anterior (e) views. f–h, Restorations of palatoquadrates showing the spiracular chamber (shaded) for Panderichthys (f), Gogonasus (g) and Eusthenopteron (h); panels f and h are based on ref. 9. ET, extratemporal bone; PO, postorbital; PP, postparietal; Spir, spiracular opening; Sp. lam., down-turned lamina of tabular for spiracular chamber; Sq, squamosal; ST, supratemporal; Ta, tabular. All bones whitened with ammonium chloride. - Long JA et al "An exceptional Devonian fish from Australia sheds light on tetrapod origins" Nature (2006) 444:199-202
a, Skull in lateral view. b, c, skull in dorsal view. d, e, X-ray micro-tomograms showing the spiracular region in lateral (d) and anterior (e) views. f–h, Restorations of palatoquadrates showing the spiracular chamber (shaded) for Panderichthys (f), Gogonasus (g) and Eusthenopteron (h); panels f and h are based on ref. 9. ET, extratemporal bone; PO, postorbital; PP, postparietal; Spir, spiracular opening; Sp. lam., down-turned lamina of tabular for spiracular chamber; Sq, squamosal; ST, supratemporal; Ta, tabular. All bones whitened with ammonium chloride. - Long JA et al "An exceptional Devonian fish from Australia sheds light on tetrapod origins" Nature (2006) 444:199-202

FIGURE 2. Pectoral fin skeleton of Gogonasus andrewsae, NMV P221807.
a–d, Right humerus in dorsal (a), mesial (b), proximal (c) and distal (d) views. e, Right ulnare in ventral view. f, Left humerus, ulna and radius articulated in dorsal view. g, Reconstructed left pectoral fin in dorsolateral view. h, Left pectoral distal elements. i, Key to h. A4, A5, distal mesomeres of the pectoral fin; Cap. hum., caput humeri; Ent. for., entepicondylar foramen; Ent. pr., entepicondyle; Pr. ven., ventral process on entepicondyle; Rad., presumed radial element; Rad. art., radial articulation; Rad. pr., radial process; Ul. pr., postaxial process of ulnare. All (except g) whitened with ammonium chloride. Scale bar, 1 cm for a–d, f–i; 0.5 cm for e. - Long JA et al "An exceptional Devonian fish from Australia sheds light on tetrapod origins" Nature (2006) 444:199-202
a–d, Right humerus in dorsal (a), mesial (b), proximal (c) and distal (d) views. e, Right ulnare in ventral view. f, Left humerus, ulna and radius articulated in dorsal view. g, Reconstructed left pectoral fin in dorsolateral view. h, Left pectoral distal elements. i, Key to h. A4, A5, distal mesomeres of the pectoral fin; Cap. hum., caput humeri; Ent. for., entepicondylar foramen; Ent. pr., entepicondyle; Pr. ven., ventral process on entepicondyle; Rad., presumed radial element; Rad. art., radial articulation; Rad. pr., radial process; Ul. pr., postaxial process of ulnare. All (except g) whitened with ammonium chloride. Scale bar, 1 cm for a–d, f–i; 0.5 cm for e. - Long JA et al "An exceptional Devonian fish from Australia sheds light on tetrapod origins" Nature (2006) 444:199-202
Panderichthys: late Devonian fish [10] roughly
contemporary or a little earlier than Eusthenopteron. Its transitional features
include:
- Loss of all midline fins
- Flatter skull with a longer snout and larger, more dorsally placed eye sockets.
- Humerus and shoulder girdle larger and more like that of a tetrapod as compared to Eusthenopteron.
- Spiracular cleft is larger than that in Eusthenopteron, and approximates that of a tetrapod. In fact, it may have been open to the outside, which is a feature associated with air breathing.
- The hyomandibular bone (homologous to the stapes, one of the middle ear bones in tetrapods) is shorter than Eusthenopteron, but still connected to the opercular bones


Figure 1 Pictures and drawings of Panderichthys rhombolepis, specimen GIT434-1.
a, Outline of the body of Panderichthys. Grey shading indicates preserved portions of Panderichthys rhombolepis specimen GIT 434-1. Redrawn from ref. 14. b, Panderichthys rhombolepis specimen GIT 434-1 with head (h) and body (b) outlined. The pelvic girdle and fin are shaded in orange. c, Pelvic girdle and fin. The matrix is distinguished from the fossil by an overlay of grey shading. d, Specimen drawing. F, femur; Fi, fibula; Fre, fibulare; Int, intermedium (proximal end of the); Pel, pelvic girdle; T, tibia. Vertical hatching indicates broken bone; grey shading indicates matrix; circles indicate thin dermal bone covering. e, Reconstruction of the pelvic fin. Thick outline indicates preserved margin, thin outline indicates inferred margin, dotted lines indicate uncertain margin. Solid black scale bars, 10 mm. - Boisvert CA "The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion" Nature (2005) 438:1145-1147
One researcher points out that the anatomical details of Panderichthys shed considerable light on the evolution of tetrapod locomotion:
The morphology of the pelvic fin of Panderichthys is a combination of primitive, transitional and unique characteristics that provide crucial information about the evolution of the tetrapod hindlimb and the origins of tetrapod locomotion.
The pectoral fin is more tetrapod-like than the pelvic fin in that the shape of the humerus is intermediate between those of osteolepiforms and basal tetrapods. [11]

FIGURE 2. Comparison of pectoral and pelvic fins.
Pectoral (a, c, e) and pelvic fins (b, d, f) of Eusthenopteron (a, b), Panderichthys (c, d) and Acanthostega (e, f) all in ventral view. F, femur; Fi, fibula; Fre, fibulare; H, humerus; Int, intermedium; R, radius; T, tibia; U, ulna; Ure, ulnare. Thick outline indicates preserved margin; thin outline indicates inferred margin; dotted lines indicate uncertain margin. Scale bars, 10 mm. Panels a and b are redrawn from ref. 3, c is from ref. 11, scale bar is from ref. 5, and e and f are redrawn from ref. 6. - Boisvert CA "The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion" Nature (2005) 438:1145-1147
Elpistostege: late Devonian lobe-finned
fish dated around 385-380 million years ago that differs from Panderichthys in
having longer snout and smaller, more rounded orbits. It resembles tetrapods in
having:
- Dorsoventrally flattened skulls
- Supraorbital ridges or “eyebrows”
- External nostrils close to the jaw margin


Source
Per Ahlberg, one of the leading experts in early tetrapod evolution notes that this fossil is a grade intermediate [12] between Panderichthys and tetrapods.
Tiktaalik: late Devonian fish [13] which is similar to Panderichthys but is even more like tetrapods, to the point that is unofficially referred to as a ‘fishapod’:

Source: Wikipedia
- Longer snout with larger eyes
- Shoulder girdle not in contact with head. This allows greater head mobility to permit raising and lowering with far more ease than other fish
- Hyomandibular bone is shorter than in Panderichthys indicating changes to the gill region and breathing mechanism
- Spiracular chamber opening is more rounded.

a, Left lateral view; b, dorsal view with enlargement of scales; and c, ventral view with enlargement of anterior ribs. See Fig. 3 for labelled drawing of skull in dorsal view. Abbreviations: an, anocleithrum; bb, basibranchial; co, coracoid; clav, clavicle; clth, cleithrum; cbr, ceratobranchial; ent, entopterygoid; hu, humerus; lep, lepidotrichia; mand, mandible; nar, naris; or, orbit; psp, parasphenoid; ra, radius; suc, supracleithrum; ul, ulna; uln, ulnare. Scale bar equals 5 cm. - Daeschler E.B., Shubin N.H, Jenkins F. “A Devonian tetrapod-like fish and the evolution of the tetrapod body plan.” Nature. 2006; 440(7085):757-63

a, Dorsal view of body with scales removed depicting orientation of ribs as preserved in NUFV 108. b, Lateral view. The ribs are shown in dorsoventral orientation. Number of ribs is estimated from the incompletely preserved series in NUFV 108. - Daeschler E.B., Shubin N.H, Jenkins F. “A Devonian tetrapod-like fish and the evolution of the tetrapod body plan.” Nature. 2006; 440(7085):757-63
The most important feature of Tiktaalik is that it has a wrist, which takes its upper fin even closer to being classed as a limb:
"The array of joints in the distal fin is functionally similar to the multiple transverse joints that characterise the carpal, metacarpophalangeal and interphalangeal joints of the tetrapod manus. The distal endoskeleton of Tiktaalik invites direct comparisons to the wrists and digits of limbed vertebrates. The intermedium and ulnare of Tiktaalik have homologues to eponymous wrist bones of tetrapods with which they share similar positions and articular relations. In both Tiktaalik and early tetrapods, the ulnare is block-shaped and articulates with multiple radials or digits, whereas the intermedium is a simple rod. The formation of a mobile transverse joint at the distal margin of these bones in Tiktaalik presages the establishment of a functional proximal carpal joint.”
“As in the digits and phalanges in a tetrapod limb, the inter-radial joints distal to this primordial wrist are more or less transversely aligned and capable of flexion and extension. The occurrence of multiple distally facing radial rows that are capable of flexion and extension is a likely antecedent condition to the dactyly of early tetrapods. The transformation of fins to limbs, then, probably entailed the elaboration and proliferation of structures, joints and functions already present in the fins of fish such as Tiktaalik.” [14]
a, Stereo pair of left pectoral fin of NUFV 108 in dorsal view showing disparity in size and position of anterior (alp) and posterior (plp) unjointed lepidotrichia and the relative position of dermal girdle elements. b, Right pectoral fin of NUFV 110 in anterior view showing preservation of anterior lepidotrichia (alp), clavicle (cl), scales (sc) and endochondral bones in articulation (H, humerus; U, ulna; u, ulnare; r, radials). The anterior lepidotrichia terminate at the elbow, thus allowing a full range of flexion at that joint. c, Right pectoral fin of NUFV 110 in ventral view showing positions of coracoid (co) and endochondral and dermal fin elements. an, anocleithrum; cb, ceratobranchial; clth, cleithrum; int, intermedium; ri, rib; suc, supracleithrum. - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006;440(7085):764-71.
a, Dorsal view; b, ventral view. Elements with stipple shading were preserved in articulation in NUFV 109 and prepared in the round. Elements with a dashed outline are reconstructed based on their presence in the articulated distal fin of NUFV 110. It is not known how many radials lie distal to the first, second and fourth in the proximal series. Note the dorsal expansion of the distal articular facets on the ulnare and third distal radial/mesomere. The dorsal expansion of these facets would have facilitated extension of the distal fin. - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006;440(7085):764-71.
a, Dorsal view; b, ventral view; c, medial view; d, posterior view. ac gle, anterior cam of glenoid facet; cf, coracoid foramen; clth, cleithrum; co, coracoid; gle, glenoid facet; sp, scapular process. The hatched area is covered by matrix. - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006;440(7085):764-71.



To summarise the above comments from the research paper:
- The bones in the pectoral fin of Tiktaalik perform similar functions to those in the tetrapod upper limb
- Furthermore, these bones are homologous to those in the tetrapod limb
- Although Tiktaalik is a fish by definition (it has scales and the presence of rays means it has fins instead of limbs), structurally and functionally, it is clearly transitional between fish and tetrapod to the point that it blurs the transition and fully warrants the name ‘fishapod’

Unlike other tetrapodomorph fishes (1), Tiktaalik has reduced the unjointed
lepidotrichia, expanded the radials to a proximal, intermediate and distal
series, and established multiple transverse joints in the distal fin. The fin
also retains a mosaic of features seen in basal taxa. The central axis of
enlarged endochondral bones is a pattern found in basal sarcopterygians and
accords with hypotheses that a primitive fin axis is homologous to autopodial
bones of the tetrapod limb. In some features, Tiktaalik is similar to
rhizodontids such as Sauripterus. These similarities, which are probably
homoplastic, include the shape and number of radial articulations on the
ulnare, the presence of extensive and branched endochondral radials, and the
retention of unjointed lepidotrichia. Figures redrawn and modified from
Glyptolepis, Eusthenopteron, Panderichthys, Acanthostega and Tulerpeton. -
Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and
the origin of the tetrapod limb. Nature. 2006;440(7085):764-71.

a, b, Anterolateral view. c, d, Ventral view. a, c, Resting posture with the fin partially flexed at the antebrachium. In this position the radius is slightly more flexed than the ulna. b, d, Resistant contact with a firm substrate entails flexion at proximal joints and extension at distal ones. The shoulder joint is flexed by ventral muscles, including the trans-coracoid muscle. The elbow is flexed (d, arrow 1), with slight pronation of the radius (d, arrow 2) and rotation of the ulna (d, arrow 3). The transverse joints distal to the ulnare and intermedium are extended (d, arrows 4). - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006;440(7085):764-71.
As the authors note:
The pectoral skeleton of Tiktaalik is transitional between fish fins and tetrapod limbs. Comparison of the fin with those of related fish reveals that the manus is not a de novo novelty of tetrapods; rather, it was assembled in fishes over evolutionary time to meet the diverse challenges of life in the marshes of Devonian aquatic evosystems. [15]

Since the publication of the initial papers, the pelvic girdle of Tiktaalik has been discovered, one that was greatly enlarged compared to other tetrapodomorphs with fins. Shubin et al note that the
…acetabulum [hip socket] is a deeply concave socket that consists of a smooth surface internally. The socket is relatively round in shape and much deeper than the corresponding joint surface in Gooloogongia or Eusthenopteron. A semicircular embayment of unfinished bone is continuous with the cranial margin of the acetabulum and may have housed cartilage or bone that participated in the joint. In Tiktaalik, the acetabulum is situated caudally on the pelvis, like other finned tetrapodomorphs. The acetabulum of Tiktaalik would have faced more laterally than that of fish but less than tetrapods. Consequently, Tiktaalik Presents a mosaic of primitive and derived conditions: like fish, it has an acetabulum that sits at the caudal margin of the pelvis, but it is more tetrapod-like in the degree to which the acetabulum faces laterally. [16]

Type specimen (NUFV108): ventral surface of cranial block (figured in ref. 6) aligned in preserved position with ventral view of the block containing the pelvic fin. (Inset) Line diagram of lepidotrichia and preserved portions of endochondral bones of pelvic fin. f, fin; i, intermedium?; l, lepidotrichia; r, radials.

Acanthostega (Top), Tiktaalik (Middle), and Eusthenopteron (Bottom) reconstructions with pectoral and pelvic girdles (proportions were derived from refs. 10, 16, and 17 and NUFV108).

Reconstruction of the skeleton of Tiktaalik roseae, (All images from ref. 16)

Acanthostega (Top), Tiktaalik (Middle), and Eusthenopteron (Bottom) reconstructions with pectoral and pelvic girdles (proportions were derived from refs. 10, 16, and 17 and NUFV108).

Reconstruction of the skeleton of Tiktaalik roseae, (All images from ref. 16)
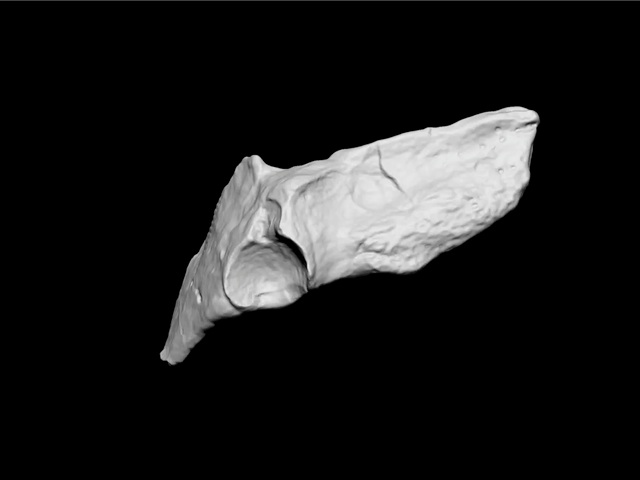
Pelvis of Tiktaalik roseae. (Source)
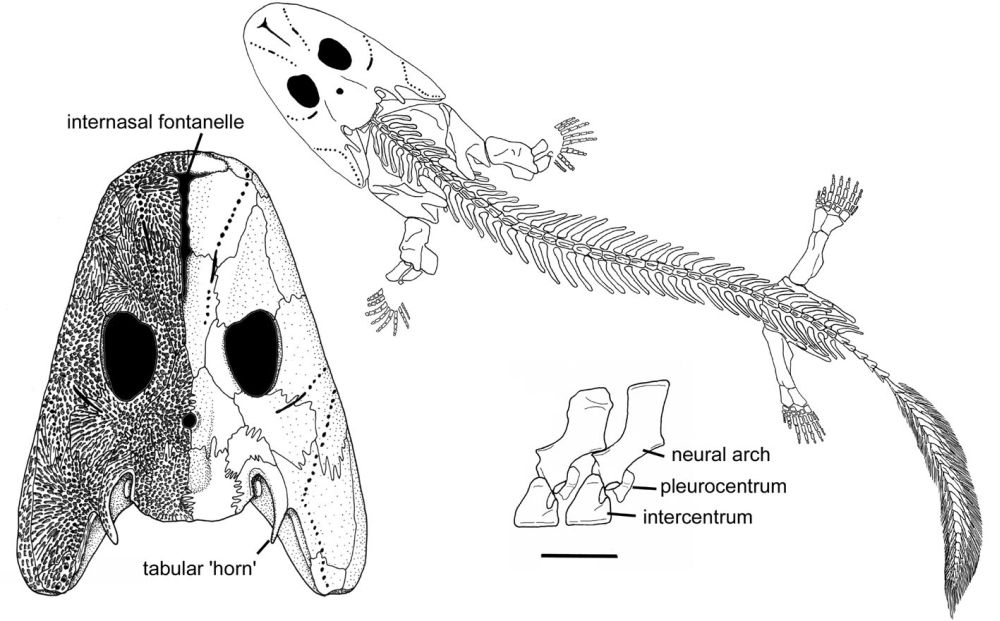
Ventastega: this is a late
Devonian tetrapod with fish-like features. [17] It is classified as a tetrapod on the basis of material from its lower jaw and
girdle.

Ventastega reconstruction. Source: Wikipedia

Ventastega reconstruction. Source: Wikipedia
Of note are:
- Its shoulder and hip girdle, which closely resemble that of the tetrapod Acanthostega, allowing one to infer the presence of digits.
- Its skull, which is intermediate in appearance between that of the fish Tiktaalik and the tetrapod Acanthostega.
- Its braincase, which shows a mix of fish-like and tetrapod-like features.

a, b, Associated skull roof (LDM G 81/775) and cheek (LDM G 81/776) in dorsal (a) and left lateral (b) views, anterior at the top. The internasal fontanelle, orbit and spiracle are indicated in a. c, d, The same specimen without the cheek in left lateral (c) and ventral (d) views, anterior at the top, showing the three-dimensionally preserved braincase. The parasphenoid and sphenoid are indicated. e, Complete lower jaw (LDM G 81/777) in medial view with coronoid fangs shown. Scale bar, 10 mm. 'LDM G' denotes the geology collections of Latvijas Dabas Muzejs, the Natural History Museum of Latvia. - Ahlberg, PE. Clack JA, Luksevics E, Blom H and Zupins I "Ventastega curonica and the origin of tetrapod morphology". (2008) Nature 453, 1199-1204 (2008)

a, Right anocleithrum (LDM G 81/778) in lateral and anterior views (from left to right), showing overlap for cleithrum. b, Left cleithrum (LDM G 81/779) and partial scapulocoracoid in lateral, anterior and mesial views (from left to right). Note the broad shallow subscapular fossa (s.fossa) and the partially preserved glenoid canal. c, Interclavicle (LDM G 81/601) in ventral view showing clavicular overlaps. d, A probable tetrapod rib (LDM G 81/781). e, Right ilium (LDM G 81/780) in anterior, lateral, mesial and dorsal views (from left to right), showing the iliac neck, dorsal process and posterior process. f, A probable tetrapod caudal fin lepidotrichium (LDM G 81/782) on a block of matrix. Scale bar, 10 mm; all specimens shown to same scale. - Ahlberg, PE. Clack JA, Luksevics E, Blom H and Zupins I "Ventastega curonica and the origin of tetrapod morphology". (2008) Nature 453, 1199-1204 (2008)

a, Whole-body reconstruction showing known skeletal elements on a body outline based on Acanthostega (modified from ref. 5; original Acanthostega body reconstruction by M. I. Coates). Scale bar, 10 cm. b, c, Skull reconstruction in lateral and dorsal views, based on material presented here and described previously21. d, Reconstructed association of skull and shoulder girdle in lateral view. e, Shoulder girdle in anterior view. Curvature of cleithrum based on LDM G 81/522 (ref. 21). Unknown bones are indicated with vertical hatching. Scale bar for b–e, 10 mm. f, g, Life reconstructions of head in lateral and dorsal views (copyright P. Renne, 2007). an, anocleithrum; ang, angular; cla, clavicle; clei, cleithrum; de, dentary; fr, frontal; icl, interclavicle; i.fon, internasal fontanelle; it, intertemporal; ju, jugal; la, lacrimal; mx, maxilla; m.ro, median rostral; na, nasal; pa, parietal; pmx, premaxilla; po, postorbital; pof, postfrontal; pop, preopercular; pospl, postsplenial; pp, postparietal; prf, prefrontal; pter, pterygoid; qj, quadratojugal; sang, surangular; scapcor, scapulocoracoid; spl, splenial; sq, squamosal; ta, tabular. - Ahlberg, PE. Clack JA, Luksevics E, Blom H and Zupins I "Ventastega curonica and the origin of tetrapod morphology". (2008) Nature 453, 1199-1204 (2008)
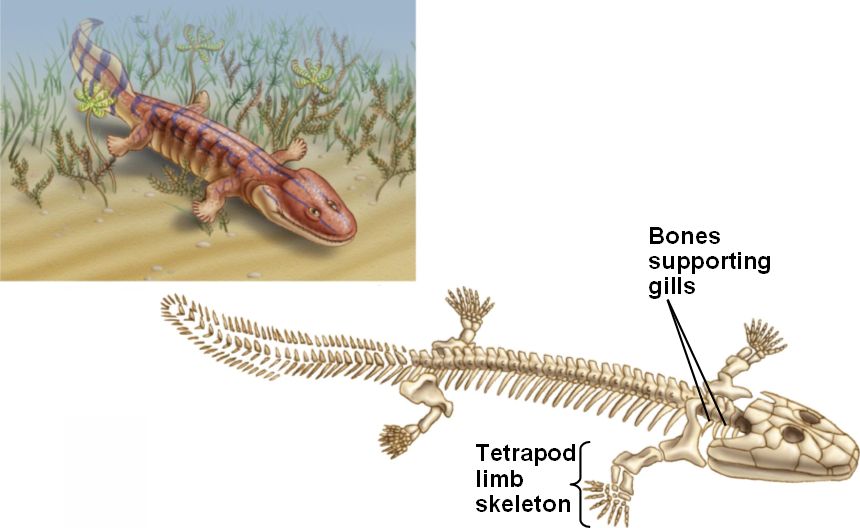
Acanthostega: was a Devonian
tetrapod living around 370-360 million years ago. It had limbs with digits, but
retained a number of fish-like characteristics. Despite the presence of legs,
it was poorly suited to terrestrial life as it could not support its weight on
land, and was therefore mainly aquatic.

Acanthostega reconstruction. Source: Wikipedia
Fish-like features included:

Acanthostega reconstruction. Source: Wikipedia
Fish-like features included:
- Gill bars
- No true elbows, knees, wrists or ankles
- Long fin rays around tail
Source
Source
Of course it was a tetrapod as it had four limbs with digits. Other specific tetrapod features [18] included:
- Head not joined to shoulder girdle
- Ear bone fitted into braincase fenestra
- Large pelvic girdle and sacral rib
- Femur and humerus were of similar size
- Tibia and fibula attached to ankle bones
- Digits on each limb

Pectoral endoskeleton of tetrapodomorph fish and a tetrapod in ventral view. Pectoral fins of: a, Eusthenopteron; b, Panderichthys; and c, Tiktaalik. d, Limb of Acanthostega. H, humerus; Int, intermedium; R, radius; U, ulna; Ure, ulnare. Scale bar, 1 cm. - Nature (2008) 456:636-638

Source
Clack is worth quoting at length to show why Acanthostega is regarded as an excellent transitional fossil:
The fish-like features of Acanthostega include a tail fin supported by long bony rays, to make an oar shape. Fin rays had been found in the tail of Ichthyostega, but they were much shorter and less numerous. The ribs of Acanthostega are short and slender and much the same all down the length of its body… Ichthyostega was known to have had broad overlapping ribs at least in its trunk region. In the skull, Acanthostega has a slot running down the midline of the snout, similar to the gap found there in Ventastega...Acanthostega also retains a set of well-ossified grooved gill bars, suggesting that these were still actively involved in gill breathing, though it probably breathed air by gulping as well.
In other respects, Acanthostega showed some typical tetrapod features. It has a robust pelvic girdle and a large femur. It is the pelvic region and the hind limb that most conspicuously distinguish a fish from a tetrapod, and that distinction is still valid among modern animals. In fish, the pelvic girdle is usually small and not attached to the vertebral column and the pelvic fin is diminutive, whereas in tetrapods, the pelvic girdle is large, attached to the vertebral column and bears the substantial hind limbs that provide most of the propulsive power in walking. In Acanthostega, although the pelvic girdle is enlarged, it is still relatively small compared with later tetrapods and was only attached to the column by soft tissue rather than a bony junction.
Some excellent specimens showed that it had eight digits on the forearm…and almost certainly eight (or maybe more) on the hind limb. That they are so neatly laid out in the fossil may be because they were enveloped in a web of skin. The wrist bones were not ossified, but it is clear that the digits were arranged in a broad arc around where those bones would have been, but the result would not have made a flexible or supportive wrist. The limb was in effect a paddle…Similarly, although the ankle bones are well ossified, they do not show an obvious joint surface along which the limb could bend. Again, the limb seems to have been a paddle. From that evidence, it was suggested that limbs with digits first evolved not for walking on land but for swimming or wading through water.
In the braincase and ear region, Acanthostega showed features in which it was distinctively more tetrapod-like than fish-like. In tetrapods, rather than a hyomandibula pivoting on the braincase wall, essentially the same bone but now termed the stapes, fits into a hole in the braincase wall called the fenestra vestibuli or fenestra ovalis. Acanthostega shows this pattern in a very early form. The arrangement later became modified into the combination of a stapes and a fenestra ovalis that is characteristic of all tetrapods and is still part of the hearing mechanism in humans. [19]
Ichthyostega: this was another
Devonian tetrapod with fish-like features, and like Acanthostega not fully
suited to terrestrial life.

Ichthyostega reconstruction. Source: Wikipedia

Source (Reconstruction derived from research by Per Ahlberg)

Ichthyostega reconstruction. Source: Wikipedia
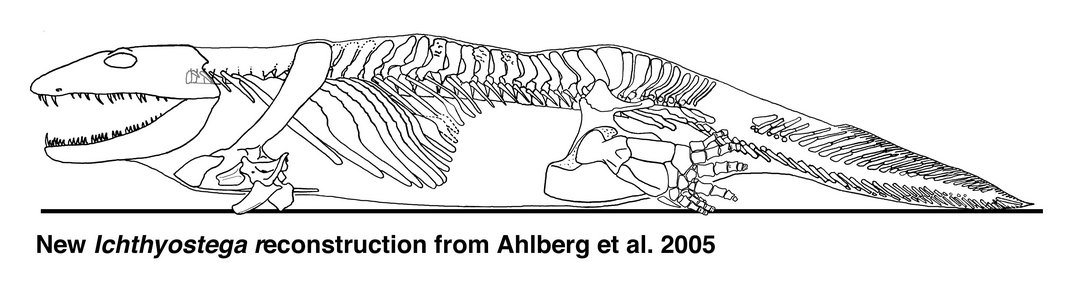
Source (Reconstruction derived from research by Per Ahlberg)

Skull roof of Ichthyostega. MGUH VP 6064 Geological Museum, Copenhagen. Photo J. A. Clack. Copyright © 1997 University Museum of Zoology, Cambridge. (http://tolweb.org/tr...kull_fossil.JPG)
Clack again summarises its status as a key transitional fossil:
If Acanthostega is a mixture of fish-like and tetrapod-like characters, Ichthyostega is a mixture of apparently aquatic specializations with others that appear more modified for land excursions. Recent study has revealed a rather different animal from the old image of an overlarge salamander-like body form.
One of the parts of the anatomy that had puzzled earlier workers was the ear region. Using newly collected and prepared fossil material, together with microcomputed tomography scanning of key fossils, the unique arrangement found in Ichthyostega has been interpreted as a highly specialized underwater auditory organ…By contrast, work on its postcranial skeleton has suggested that it may have had a unique form of locomotion on land. The vertebral column is differentiated into separate regions, including a lumbar portion that seems to have accommodated dorsoventral flexion The shoulder girdle and forearm are large and robust, suggesting extensive musculature, perhaps for pulling the animal along on land. Its hind limb is quite similar in construction to that of Acanthostega, that is to say a paddle with no obvious ankle joint, and it has seven toes arranged in a unique pattern. A group of three small toes form a strengthening bar along the leading edge, with four stouter ones behind. It is likely that, as in Acanthostega, these were contained in a web of skin. The hind limb may have given stability and purchase on the substrate and been used as a flipper in water, but it was not a conventional walking leg. It is no surprise that this peculiar morphology caused problems for early workers trying to understand Devonian tetrapods. It shows that even by that time, tetrapods had diversified into a wider range of morphologies than was previously appreciated. Analysis of all the skull specimens of Ichthyostega has shown changes to proportions and skull bone ornamentation that varies throughout its history and that document microevolution at the specific level in these very early tetrapods. Skulls from lower formations are narrower, with finer ornament, than those from higher up in the geological sequence. [20]

Hindlimb of Ichthyostega; specimen collected in 1987. Geological Museum, Copenhagen MGUH Field number 1349. Photo M. Coates. Copyright © 1997 University Museum of Zoology, Cambridge. (http://tolweb.org/tr...a_hind_limb.JPG)
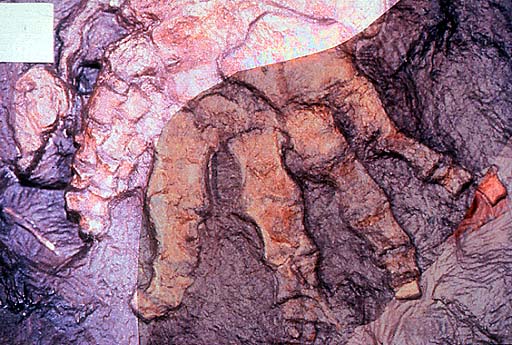
Photographs of part and counterpart superimposed to show seven digits (specimen collected in 1987). Geological Museum, Copenhagen MGUH Field number 1349. Photo M. Coates. Copyright © 1997 University Museum of Zoology, Cambridge. (http://tolweb.org/tr...ostega_foot.JPG)
Pederpes: this was a tetrapod
living around 360 to 350 million years ago that had five digits on its hind
limbs, and six digits – five formed and one vestigial – on its front limbs.
This details more evolutionary change from the earlier tetrapods as inferred
from Ichthyostega and Acanthostega, which had more than five digits.

Source: Wikipedia
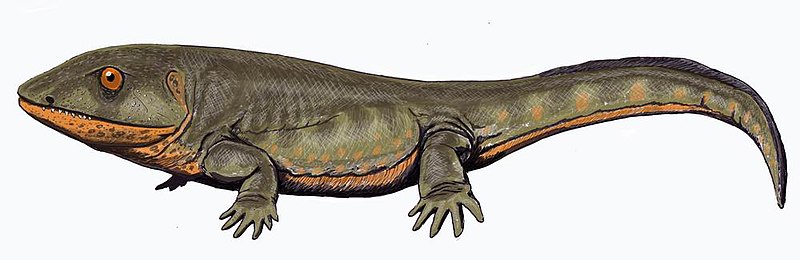
Source: Wikipedia
Clack notes:
“This animal, belonging to a group known as the whatcheeriids, not only shows some primitive features in which it resembles the Devonian forms but also has more advanced features that suggest the beginnings of terrestriality. Its hind limb was fairly typical of a conventional tetrapod from the Late Carboniferous and appears to have had five digits. The foot appears to have pointed forward as in a terrestrially walking animal, rather than to the side as in Acanthostega. However, the forelimb may have had more than five digits, because the only two that are known are both extremely small and resemble the supernumeraries of Acanthostega and Ichthyostega. Pederpes has broad flanges on its ribs that resemble those of Ichthyostega in some ways. Its stapes is like that of Acanthostega.” [21]

a, GLAHMS 100815 Pederpes finneyae, photograph of holotype and only specimen. Scale bar, 100 mm; b, Map of the holotype, showing elements preserved on the reverse in grey. Scale bar, 10 mm. Fro, frontal; intemp, intertemporal; jug, jugal; lac, lacrimal; max, maxilla; nas, nasal; pmx, premaxilla; po, postorbital; pofr, postfrontal; prefr, prefrontal; preop, preopercular; qj, quadratojugal; scapcor, scapulocoracoid. - Clack J.A. "An early tetrapod from 'Romer's Gap'" Nature 418 (2002) :72-76 doi:10.1038/nature00824

Close-up of the skull of Pederpes showing the stapes. - Clack J.A. "An early tetrapod from 'Romer's Gap'" Nature 418 (2002) :72-76 doi:10.1038/nature00824

a, Humerus, radius, ulna and manual digits from the right forelimb. The minute irregular elements at the base of the tiny digit might be remains of metacarpals. b, Pedal digits and femur from the right hindlimb, shown reversed. c, Reconstructions of manual (left) and pedal digits (right) of Pederpes to the same scale, showing tiny manual digit and asymmetrical metatarsals respectively. Scale bar, 10 mm. In Pederpes, the phalanges of the tiny manual digits are about one-third the length of the pedal phalanges, whereas in Acanthostega and the Carboniferous forms such as Greererpeton and Proterogyrinus, manual phalanges are about one-half the length of the pedal phalanges. - Clack J.A. "An early tetrapod from 'Romer's Gap'" Nature 418 (2002) :72-76 doi:10.1038/nature00824
Again, we see the mosaic of
fish-like and tetrapod-like features, but with a definite progression towards
terrestrial life. This – yet again – is another transitional fossil which along
with the earlier taxa shown illustrate large-scale evolutionary change from
lobe-finned fish to tetrapods.
Conclusion
What we have seen here clearly demonstrates the large-scale evolutionary changes that took place between lobe-finned fish and tetrapods. Excellent evolutionary transitions can be found elsewhere, but space prohibits further comprehensive examination. Some of the more notable evolutionary transitions include the evolution of whales [22] from artiodactyls, the evolution of the mammalian jaw and middle ear [23-26] in which two reptile jaw bones became detached and incorporated into the mammalian middle ear as the incus and malleus, as well as the evolutionary trends in terrestrial hoofed mammals.[27]
Pearce is wrong to allege or imply there are no transitional fossils. They about in the fossil record, and document large-scale evolutionary change. Creationists such as Pearce have failed to grasp that the concept of “missing links” is not one maintained by palaeontologists, and his use of it betrays a fundamental ignorance of the science he criticises. Clack and Ahlberg make this point quite emphatically:
What we have seen here clearly demonstrates the large-scale evolutionary changes that took place between lobe-finned fish and tetrapods. Excellent evolutionary transitions can be found elsewhere, but space prohibits further comprehensive examination. Some of the more notable evolutionary transitions include the evolution of whales [22] from artiodactyls, the evolution of the mammalian jaw and middle ear [23-26] in which two reptile jaw bones became detached and incorporated into the mammalian middle ear as the incus and malleus, as well as the evolutionary trends in terrestrial hoofed mammals.[27]
Pearce is wrong to allege or imply there are no transitional fossils. They about in the fossil record, and document large-scale evolutionary change. Creationists such as Pearce have failed to grasp that the concept of “missing links” is not one maintained by palaeontologists, and his use of it betrays a fundamental ignorance of the science he criticises. Clack and Ahlberg make this point quite emphatically:
The concept of ‘missing links’ has a powerful grasp on the imagination: the rare transitional fossils that apparently capture the origins of major groups of organisms are uniquely evocative. But the concept has become freighted with unfounded notions of evolutionary ‘progress’ and with a mistaken emphasis on the single intermediate fossil as the key to understanding evolutionary transitions. Much of the importance of transitional fossils actually lies in how they resemble and differ from their nearest neighbours in the phylogenetic tree, and in the picture of change that emerges from this pattern.” [28]
The picture of change that we see is that of unambiguous large-scale evolutionary change, one which exposes as vacuous and uninformed claims by creationists such as David Pearce that transitional forms are not found in the fossil record.
[1] Prothero D “Evolution: What the Fossils Say and Why
It Happened” (2007: Columbia University Press)
[2] Pearce op cit p 4
[3] A. M.
Shedlock and S. V. Edwards. Amniotes (Amniota). Pp. 375–379 in The
Timetree of Life, S. B. Hedges and S. Kumar, Eds. (Oxford University Press,
2009)
[4] Mead LS "Transforming Our
Thinking about Transitional Forms" Evo Edu Outreach (2009) 2:310–314
[5] Zhu, M and Ahlberg P.E.“The origin of the internal nostril of tetrapods” (2004)
Nature 432, 94-97
[6] Janvier P Nature (2004) 234:23-24
[7] Watson DMS “The Internal Ear of Osteolepis” J Anat 1925 July 59(Pt
4) 385-386
[8] Clack J.A. “The Fish-Tetrapod Transition: New Fossils and Interpretations”
(2009) Evo Edu Outreach 2:213-223
[9] . Long J.A. et al An exceptional Devonian fish from Australia sheds
light on tetrapod origins Nature (2006) 444:199-202
[10] Boisvert C.A. The pelvic fin and girdle of Panderichthys and the origin of
tetrapod locomotion. Nature. 2005;438(7071):1145-7
[11] ibid, p 1145
[12] Ahlberg P.E. et al “A near-tetrapod from the Baltic Middle Devonian”
Palaeontology (2000) 43(3);533-548
[13] Daeschler E.B., Shubin N.H, Jenkins F. “A Devonian tetrapod-like fish and the
evolution of the tetrapod body plan.” Nature. 2006; 440(7085):757-63
[14] Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and
the origin of the tetrapod limb. Nature. 2006;440(7085):764-71.
[15] Shubin, Daeschler, Jenkins. op cit, p 769
[16] Neil H. Shubin NH, Daeschler EB, and Jenkins FA "Pelvic girdle and fin of Tiktaalik roseae" Proc Natl Acad Sci USA (2014) 111:893-899
[16] Neil H. Shubin NH, Daeschler EB, and Jenkins FA "Pelvic girdle and fin of Tiktaalik roseae" Proc Natl Acad Sci USA (2014) 111:893-899
[17] Ahlberg, PE. Clack JA, Luksevics E, Blom H and Zupins I "Ventastega
curonica and the origin of tetrapod morphology". (2008) Nature 453,
1199-1204 (2008)
[18] Clack JA. Acanthostega. A very primitive tetrapod from the Late Devonian witheight fingers and toes. A 'transitional form' between 'fishes' and 'tetrapods'.
http://www.theclacks...anthostega.html Accessed: 22nd January 2014
[19] Clack (2009), op cit. p 218-219
[20] ibid, p 219-220
[21] ibid, p 220
[22] Thewissen JGM et al “From Land to Water: the Origin of Whales,
Dolphins, and Porpoises” Evo Edu Outreach (2009) 2:272–288
[23] Meng J et al “Transitional mammalian middle ear from a new Cretaceous
Jehol eutriconodont.” (2011). Nature 472:181-185
[24] Kermack KA, Mussett F, Rigney HW (1981). "The skull of Morganucodon".
Zoological Journal of the Linnean Society 71 (1):
1–158
[25] Luo, Z., Chen, P., Li, G., & Chen, M. (2007). “A new eutriconodont mammal
and evolutionary development in early mammals.” Nature 446:288-293
[26] Romer AS “Cynodont Reptile with Incipient Mammalian Jaw Articulation” Science
(1969) 166:881-882
[27] Prothero DR "Evolutionary Transitions in the Fossil Record of Terrestrial
Hoofed Mammals" Evo Edu Outreach (2009) 2:289–302
[29] Ahlberg
PE, Clack JA “A Firm Step From Water to Land” Nature (2006) 440:747-749


Comments
Post a Comment