Human / chimpanzee common ancestry - the current fossil evidence - Part 1
Christadelphian attacks [1] on the evidence for human evolution are
hopelessly dated, relying in general on such special creationist staples
as Nebraska Man, Piltdown man and other long-rebutted [2] assertions. Another is the claim that the amount of hominid
fossils could barely fit on a billiard table, a claim which betrays considerable ignorance of the subject of human evolution:
Opponents of scientific biology are fond of dismissing that record as a pathetic handful of controversial fragments. If that were so, this book would be a lot shorter. An often-repeated creationist canard insists that all known human fossils would fit on a billiard table. This was probably true in the late 19th century but it has not been true for a hundred years. Known human fossils number in the thousands and represent the remains of hundreds of individuals. They are more numerous and better-studied than the fossils of any comparable vertebrate group, because the intense interest that people have in the bones of their ancestors has driven them to devote far more effort to collecting and studying fossil humans than (say) fossil horses or herring. Having seen most of the major collections of human fossils in the world’s museums, we can assure our readers that those collections can no longer be laid out on a billiard table. It would be hard to cram them all into a boxcar. [3]
The biggest misunderstanding however concerns the search for the
'missing link', a phrase which betrays a complete misunderstanding of
how evolutionary biologists think. When special creationists refer to a
'missing link', they are thinking in terms of evolution as a ladder,
with humans at the top, and 'lower animals' below. That is false.
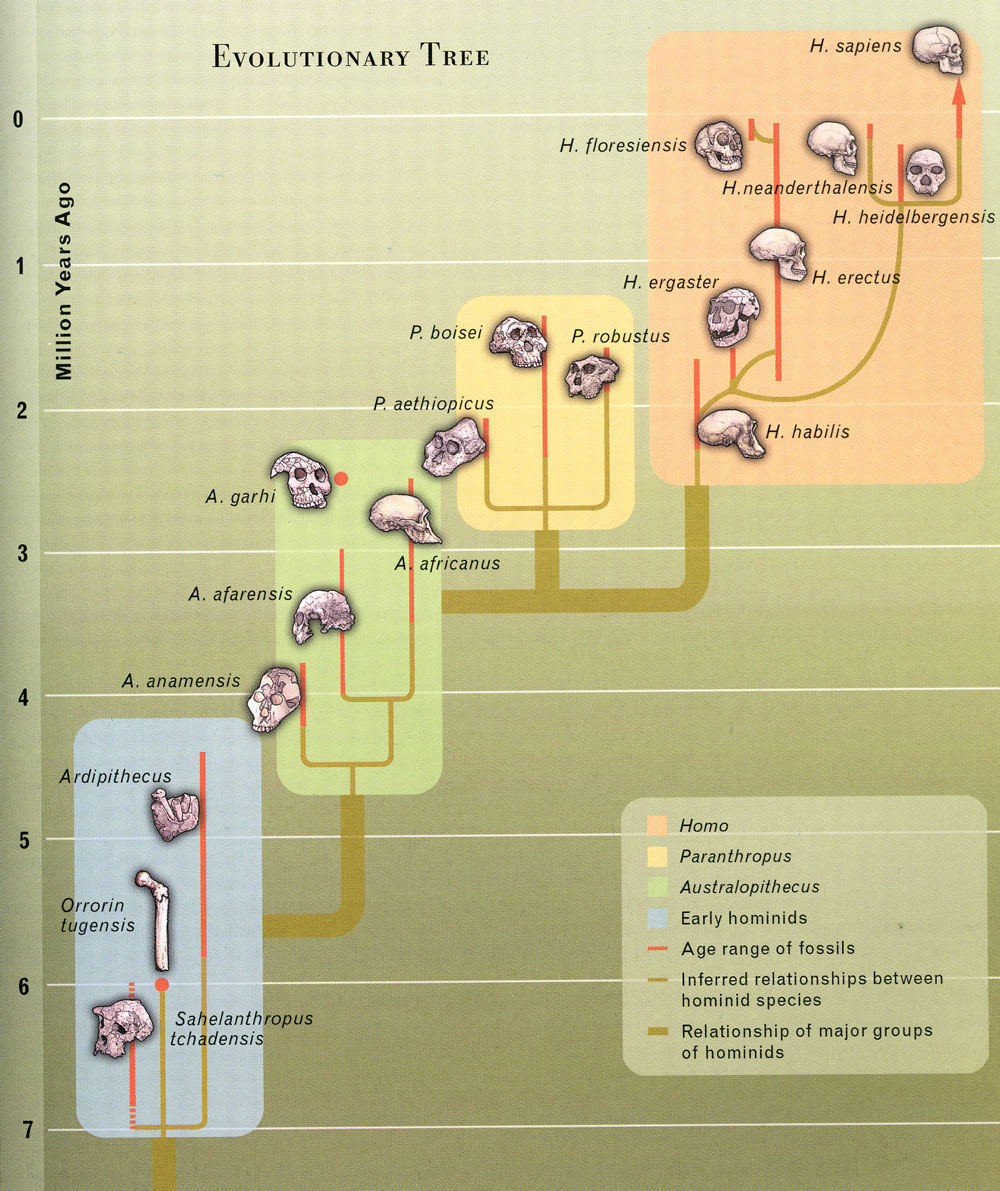
Rather, they should be thinking in terms of a tree:

The term 'missing link' is inaccurate as it presupposes evolution along a linear pathway. Rather, given that evolution is a branching process, what palaeoanthropologists (and palaeontologists in general) are looking for are relationships between fossils that allow them to construct an evolutionary family tree. Given the
fragmentary fossil record, it is no small miracle that we have three
fossil species from around that time, which provide insight into what
the common ancestor of humans and chimpanzees may have looked like. From there on, the fossil
record of human evolution becomes far more rich. At the very least, it
dispels the special creationist myth that there is no convincing fossil
evidence for human evolution.
Fossils near the base of the human evolutionary tree
Currently, we have three fossils close to the human-chimpanzee split:
Orrorin tugenensis
Sahelanthropus tchadensis
Ardipithecus ramidus
This post will look at Orrorin tugenensis, with the subsequent posts looking at Sahelanthropus tchadensis and Ardipithecus ramidus. It is unlikely that any of these species represents the common ancestor of humans and chimpanzees, but as I said earlier, palaeontologists are more concerned with establishing relationships between fossils in order to recreate the evolutionary tree. We can infer much about the common ancestor even without the actual fossil.
Orrorin tugenensis
Orrorin tugenensis was a Miocene hominin found in Kenya in 2000 by
Brigitte Senut and Martin Pickford [4] that has been dated to
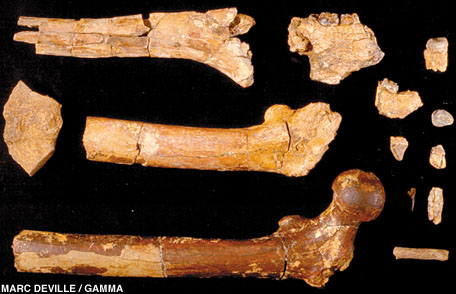
approximately 6 million years ago. The remains discovered are shown
below, followed by a description from the paper:

Hominid with jugal teeth smaller than those of Australopithecines; upper central incisor large and not shovel-shaped, with thick enamel; upper canine short with a shallow and narrow vertical mesial groove, apical height low; small triangular upper M3s; corpus mandibularis relatively deep below M3; lower P4 with offset roots and oblique crown; small Homolike rectangular lower M2 and M3s; thick enamel on lower cheek teeth; buccal notch well developed which imparts a bilobate profile to the buccal surface; no cingulum on molars. Femur with a spherical head rotated anteriorly, neck elongated and oval in section, lesser trochanter medially salient with strong muscle insertions, deep digital fossa; humerus with a vertical brachioradialis crest; proximal manual phalanx curved; dentition small relative to body size. [5]
What that means is that they found teeth, jaw fragments, a femur
and a humerus. We have no skull sadly, but we do have post-cranial data
(bones other than the skull) which is important for us to work out
whether Orrorin tugenensis was bipedal. By examining the thigh bone, we can work this out.

Morphological comparisons among femora of or attributed to (A) P. troglodytes, (B) O. tugenensis (BAR 1002'00), (C and D)Paranthropus robustus (SK 97 and SK 82, reversed), (E) A. afarensis (A.L. 288-1ap), (F) Paranthropus boisei (KNM-ER 1503, reversed), (G) early Homo (KNM-ER 1481), and (H) modern H. sapiens.
Like other early hominin femora (C to F), BAR 1002'00 (B) is distinct
from those of modern humans (H) and great apes (A) in having a long,
anteroposteriorly narrow neck and wide proximal shaft. Early Homo femora
(G) have larger heads and broader necks compared to early hominins. In
addition to these features, modern human femora (H) have short necks and
mediolaterally narrow shafts. Scale bar, 2 cm.
Source: Richmond B.G., Jungers W.L. "Orrorin tugenensis Femoral Morphology and the Evolution of Hominin Bipedalism" Science (2008) 319:1662-1665
The authors of this paper felt that Orrorin tugenensis was bipedal, as they announced:
The postcranial evidence suggests that Orrorin tugenensis was already adapted to habitual or perhaps even obligate bipedalism when on the ground, but that it was also a good climber. [6]
The authors published a family tree, showing where they felt Orrorin tugenensis was positioned:

Note that they place Orrorin tugenensis after the Australopithecine and Homo lines had diverged, close to this split.
Not everyone was convinced by this. However, further studies
performed on the bones strengthened the case for those who believed Orrorin tugenensis was bipedal.
Our results show that the internal distribution of cortical bone in its femoral neck constitutes direct evidence for frequent bipedal posture and locomotion in this Late Miocene ancestor. In known features, external and internal, BAR100200 exhibits a total morphological pattern distinct from African apes, diagnostic of bipedal locomotion, and appropriate for a population standing at the dawn of the human lineage. [7]
Did that clinch the case? Not quite. The CT scanning performed on
the bones did not have very good resolution - a recent study (2008) has
clarified the issue, and concluded that Orrorin tugenensis was quite likely bipedal, but was not more closely related to Homo than to Australopithecus:
In light of the marked changes in femoral anatomy from at least 2 Ma to the present, the close morphological similarity between femora of O. tugenensis at 6 Ma and Australopithecus and Paranthropus in the later Pliocene is especially pronounced... Although among the early hominin taxa, Orrorin has the smallest Mahalanobis D2 distance from the modern human centroid, BAR 1002’00 is much closer to all early hominin taxa than to the modern human centroid in shape space. The external morphology of O. tugenensis provides no indications of differences in bipedal gait compared to Australopithecus or Paranthropus. This suggests that the pattern of bipedal gait characteristic of australopiths evolved very early in the human lineage, and perhaps they were also the characteristics of the first bipedal hominins. This form of bipedalism appears to have persisted as a successful locomotor strategy for as long as 4 million years. Additional lower limb fossils from the late Miocene and early Pliocene will be needed to test this hypothesis.
The similarity between O. tugenensis and australopith femora weakens support for scenarios in which O. tugenesis is ancestral to Homo to the exclusion of A. afarensis. Instead, the overall primitive hominin morphology of the O. tugenensis femur, along with primitive dental anatomy, is consistent with the more parsimonious hypothesis that it is a basal member of the hominin clade. In sum, the comparative biomechanical anatomy of O. tugenensis femora suggests that O. tugenensis is a basal hominin adapted to bipedalism, and current evidence suggests that an Australopithecus-like bipedal morphology evolved early in the hominin clade and persisted successfully for most of human evolutionary history. (Emphasis mine) [8]
It would definitely be premature to declare Orrorin tugenensis directly on the line leading to Homo sapiens, but it is eminently reasonable to assume that it is a primitive hominin close to the time when the lines leading to Homo and Pan diverged.
References
1. The Creation and Evolution The Christadelphian Advocate Vol 7 No. 1
2. Creationism and Human Evolution
3. Cartmill, Matt; Smith, Fred H. (2009) The Human Lineage New Jersey, Wiley Publishing p xi
4. Senut B et al "First hominid from the Miocene (Lukeino Formation, Kenya)" Earth and Planetary Sciences (2001) 332:137–144
5. ibid, p 139
6. ibid, p 142
7. K. Galik et al "External and Internal Morphology of the BAR 1002'00 Orrorin tugenensis Femur" Science (2008) 305:1450-1453
8. Richmond BG, Jungers WL "Orrorin tugenensis Femoral Morphology and the Evolution of Hominin Bipedalism" Science (2008) 319:1662-64


{kind=link}
{kind=link}
Comments
Post a Comment