The Christadelphian magazine and evolution. Part 1 - Allan Harrison
While long overdue, the growing
recognition that the fact of evolution does not threaten any core Christadelphian
doctrine is welcome news. Regrettably, some of our magazines have chosen to
respond to this by attacking evolutionary biology, a move which is as
ill-advised as the decision by the Catholic Church to attack Galileo and
heliocentrism. In particular, since the start of this year The Christadelphian
has commenced a series of articles aimed at defending the “historicity and
literality of the Genesis record”, and announced that after this, they will
examine “some
of the doctrinal implications arising from our belief in these early chapters.”
[1]
Given that no fundamental Christadelphian doctrine is affected by an evolutionary origin of the human race, [2] it is hard to see why The Christadelphian is seeking to tie orthodoxy to special creationism. More to the point, such a move is unwise at best, given the risk of destroying the faith of Christadelphian special creationists who buy that argument, then discover the weight of evidence against special creationism, and subsequently feel they have lost the foundation of their belief.
Given that no fundamental Christadelphian doctrine is affected by an evolutionary origin of the human race, [2] it is hard to see why The Christadelphian is seeking to tie orthodoxy to special creationism. More to the point, such a move is unwise at best, given the risk of destroying the faith of Christadelphian special creationists who buy that argument, then discover the weight of evidence against special creationism, and subsequently feel they have lost the foundation of their belief.
The first article [3] which specifically
argues from the science is from Allan Harrison, a retired science educator [4]
whose attack on evolution is ultimately an argument from personal incredulity,
a conflation of abiogenesis and evolutionary biology, and a regrettable failure
to properly quote the scientists whose views he enlists to bolster his argument.
Harrison begins by asserting that
“[t]he discovery of DNA, containing all the precise information required for
life, points to an intelligent mind at the outset. Alternatives proposed by
evolutionists rely on random occurrences, the probabilities of which are impossibly
small.” Assertion is not proof, and Harrison not only neglects to substantiate his
claim, but makes the common special creationist error of arguing that since the
odds of a protein forming by chance is so remote as to be astronomical,
abiogenesis is impossible.
While there is no universally-accepted theory of abiogenesis, Harrison’s appeal to huge probabilities betrays a considerable lack of understanding of the basic scientific principles, and is therefore fallacious.
While there is no universally-accepted theory of abiogenesis, Harrison’s appeal to huge probabilities betrays a considerable lack of understanding of the basic scientific principles, and is therefore fallacious.
Molecular pharmacologist Ian Musgrave neatly
skewers the argument Harrison and other
special creationists invoke:
So the calculation goes that the probability of forming a given 300 amino acid long protein (say an enzyme like carboxypeptidase) randomly is (1/20)^300 or 1 chance in 2.04 x 10^390, which is astoundingly, mind-beggaringly improbable. This is then cranked up by adding on the probabilities of generating 400 or so similar enzymes until a figure is reached that is so huge that merely contemplating it causes your brain to dribble out your ears. This gives the impression that the formation of even the smallest organism seems totally impossible. However, this is completely incorrect.Firstly, the formation of biological polymers from monomers is a function of the laws of chemistry and biochemistry, and these are decidedly not random.Secondly, the entire premise is incorrect to start off with, because in modern abiogenesis theories the first "living things" would be much simpler, not even a protobacteria, or a preprotobacteria (what Oparin called a protobiont and Woese calls a progenote), but one or more simple molecules probably not more than 30-40 subunits long. These simple molecules then slowly evolved into more cooperative self-replicating systems, then finally into simple organisms. An illustration comparing a hypothetical protobiont and a modern bacteria is given below
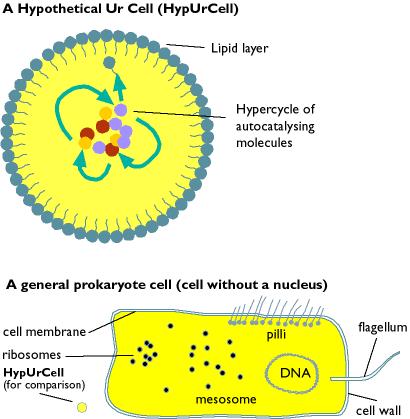
Making such an elementary mistake in the
preface of his article hardly bodes well.
Creation and Divine
Agency
It is ironic, given how some special
creationists are furiously trying to argue against the position that God has
revealed himself both in the book of nature as well as the Bible, that Harrison,
in asserting that the genome is “our Maker’s imprint”, uses language strikingly
similar to the two-book view in his opening paragraphs:
As we wait for Jesus Christ, we thank God for His wonderful gift of information in the Bible and His creation. [6]
Furthermore, in citing Romans 1:20, Harrison declares that “deniers of His creative genius
are said to be ‘without excuse’”. Evolutionary creationists yield to no one in
their appreciation of the Mind behind the universe, but they recognise two
points that special creationists fail to grasp:
- References to divine creation do not specify the mode of creation, which means that it is illegitimate to assume that references to creation are automatically special creation. The concept of divine agency, in which God is said to have achieved something despite using secondary causes, is well-established [7], and failing to appreciate this simple fact lies behind the special creationist mistake of conflating intention with mechanism.
- A diligent, honest examination of the ‘gift of information’ in creation not only shows unarguable evidence for common descent.
Much of the first page of Harrison’s article
labours under the mistake that collecting a shopping list of references to
creation (Isa 45:18, Psa 33:6-9, Psa 104:24, Acts 17:29) somehow proves special
creation.
Given the reality of Divine agency, where God is described as working through secondary causes, with ultimate authority and causality assigned to him, it is insufficient merely to cite a verse as ‘irrefutable’ evidence for special creation, as those verses say nothing about agency, but ultimately final cause.
One could just as readily cite Matthew 5:45 to argue against planetary motion and atmospheric physics, or Jeremiah 1:5 to argue against developmental biology. Failure to recognise agency is one of the main problems with special creationism and the naïve, literal reading of the Bible which underlies it.
Given the reality of Divine agency, where God is described as working through secondary causes, with ultimate authority and causality assigned to him, it is insufficient merely to cite a verse as ‘irrefutable’ evidence for special creation, as those verses say nothing about agency, but ultimately final cause.
One could just as readily cite Matthew 5:45 to argue against planetary motion and atmospheric physics, or Jeremiah 1:5 to argue against developmental biology. Failure to recognise agency is one of the main problems with special creationism and the naïve, literal reading of the Bible which underlies it.
For special creationists, the ‘wonderful gift
of information’ in creation, when analysed, is a double-edged sword, as such
analysis argues powerfully against special creation. The highly respected
evolutionary biologist John Avise, in a 2010 PNAS paper notes that:
“…many complex biological traits are gratuitously complicated, function poorly, and debilitate their bearers. Furthermore, such dysfunctional traits abound not only in the phenotypes but inside the genomes of eukaryotic species. Here, I highlight several outlandish features of the human genome that defy notions of ID by a caring cognitive agent. These range from de novo mutational glitches that collectively kill or maim countless individuals (including embryos and fetuses) to pervasive architectural flaws (including pseudogenes, parasitic mobile elements, and needlessly baroque regulatory pathways) that are endogenous in every human genome. Gross imperfection at the molecular level presents a conundrum for the traditional paradigms of natural theology as well as for recent assertions of ID, but it is consistent with the notion of nonsentient contrivance by evolutionary forces.” [8]
None of this detracts from the fact that
random mutation and natural selection together form a powerful creative engine
capable of considerable design elegance, a fact exploited by scientists and
engineers who use evolutionary algorithms capable of design far more elegant
than intelligent designers are able to achieve [9]. However, evolution is
blind, unable to look ahead further than what will achieve the immediate goal,
which means that the footprint of historical contingency can be seen in
suboptimal design, as Avise and others have pointed out. [10].
The parallel with human history and natural
history is instructive. It is axiomatic for believers that history is not just
one thing happening after another, but something that is working towards a
divine goal. God however works through human beings, whose free will means they
are capable of causing considerable suffering and evil.
Likewise, God has been working through natural history to achieve a final goal – a form of life capable of bearing his image, but his utilisation of evolutionary mechanisms means that suboptimality and flawed design are inevitable. Attacking evolution for suboptimal design and natural suffering makes as little sense as denying that God works in human history because of the Rwandan, Cambodian, and Jewish genocides of the 20th century.
Likewise, God has been working through natural history to achieve a final goal – a form of life capable of bearing his image, but his utilisation of evolutionary mechanisms means that suboptimality and flawed design are inevitable. Attacking evolution for suboptimal design and natural suffering makes as little sense as denying that God works in human history because of the Rwandan, Cambodian, and Jewish genocides of the 20th century.
Conflating abiogenesis
and evolution
Two of the fundamental errors made by special
creationists are the failure to properly define evolution, and the conflation
of abiogenesis with evolutionary biology. This confusion means that problems -
real or imagined – either with abiogenesis or the currently accepted mechanism
of evolutionary change are treated by the special creationist as somehow
falsifying the overwhelming evidence for common descent and large-scale
evolutionary change. [11] Harrison makes this
error early in his article, and it completely undermines his argument.
Harrison claims that the “biggest
problem for evolution” is “the information that made life possible”. This is
false, if only because he fails to define what
he means by evolution. Is he referring to evolutionary biology? If so, then he
is flat-out wrong because the origin of biological information poses no problem
for evolutionary biology. Point mutation, gene duplication., gene fusion, whole
genome duplication, and horizontal gene transfer are all biological mechanisms
that provide the raw material for evolution, and therefore the source of
information in the genome. The evolutionary biologist and biology information
theory expert Tom Schneider pointed out over 15 years ago how biological
information can evolve:
How do genetic systems gain information by evolutionary processes? Answering this question precisely requires a robust, quantitative measure of information. Fortunately, 50 years ago Claude Shannon defined information as a decrease in the uncertainty of a receiver. For molecular systems, uncertainty is closely related to entropy and hence has clear connections to the Second Law of Thermodynamics. These aspects of information theory have allowed the development of a straightforward and practical method of measuring information in genetic control systems. Here this method is used to observe information gain in the binding sites for an artificial ‘protein’ in a computer simulation of evolution. The simulation begins with zero information and, as in naturally occurring genetic systems, the information measured in the fully evolved binding sites is close to that needed to locate the sites in the genome. The transition is rapid, demonstrating that information gain can occur by punctuated equilibrium. (Emphasis mine) [11]
Given that
Schneider’s paper is well over a decade old, it is staggering that Harrison fails either to cite it, or provide a
substantive response to it. Either way, Harrison’s
claim that the origin of information provides a major problem for evolution, if
by evolution he is not referring to abiogenesis, is false.
Even if Harrison is referring to abiogenesis,
the origin of life from non-life, his argument does not provide a knock-down
argument against evolution, simply because the fact of common descent remains
irrespective of whether we have a robust theory of abiogenesis or not. Solid
state physics and general relativity do not collapse simply because we cannot
unite quantum physics and general relativity.
If Harrison is referring to abiogenesis, then central to his assertion is the claim that the genetic code could not evolve. Again, while we have no robust theory of abiogenesis, and as biochemist Larry Moran notes, we do not know at the moment how life began [12], this does not mean that we are completely in the dark about how the genetic code could have evolved. Astrobiologist Stephen Freeland notes that:
First, it has been discovered that the genetic code is not universal. Around a dozen or so minor variations exist. These variations are mostly codes in which one or more genetic codons have altered their amino acid “meanings.” Some involve a more significant change – the addition of a 21st or 22nd amino acid. Everything indicates that these genetic codes evolved from the standard genetic code during the past few hundred million years, and continues to evolve today. Arguments for the evolvability of the code are strengthened by the finding that amino acids are assigned to genetic code-words non-randomly. In particular, codons are assigned to amino acids in such a pattern that common mutations produce minor variations as proteins are decoded. A growing body of evidence connects this feature of the code to the idea that considerable evolution by natural selection had gone into shaping this system. Everything suggests that the genetic code is evolved and evolvable after all.The second major insight into the origins of genetic coding is that multiple, independent lines of evidence suggest the standard amino acid alphabet of 20 building-blocks grew from a smaller earlier alphabet corresponding to an earlier stage in genetic code evolution. Many variations have been proposed. Most derive their views by considering only one or two types of evidence; sophisticated calculations of the amino acid sequences of truly ancient proteins, the repertoire of amino acids found in meteorites; simulations of an early, pre-biological planet Earth and so on. What is interesting is an un-looked for match between the broad findings of these different approaches. In particular, different approaches end up dividing the 20 amino acids of modern organisms into 10 that were around in the earliest systems, and 10 that arrived later, as by-products of early biological evolution. The members of each group are remarkably consistent, hinting directly at the process by which the genetic code evolved, growing more complex over time from simpler beginnings. Recent findings are also starting to make sense of why natural selection created this particular alphabet of building blocks.The third line of insight takes us backwards to the possible origins of genetic coding. Some scientists have used the SELEX approach described in a companion paper by Watts to define mini-sequences of RNA that specifically bind to a particular amino acid. Although results have been patchy, some amino acids seem to associate with surprising choosiness to the code-words assigned to them in the standard genetic code. This association suggests that the earliest steps in genetic coding may have been nothing more than simple physical affinities between two types of chemical.Between them, these insights represent significant progress from the impossibly self-referential system viewed by Crick and those around him just 50 years ago. This half-century of research indicates that the standard genetic code at work in modern cells may be a product of substantial evolution that had taken place by around 3 billion years ago. [13]
Irrespective of whether Harrison
was referring to evolutionary biology or abiogenesis, his assertion about
information is totally incorrect (or at best, considerably overstated).
Harrison’s failure to properly define evolution appears to be corrected a little later when he asserts:
“Darwinian evolution, despite its popularity, lacks the analytical support enjoyed by the theories of true science.”
But as the rest of his article completely
fails to address the modern synthetic theory of evolution, and instead
concentrates on a superficial, and poorly informed creationist attack on
abiogenesis, Harrison has mislead his readers by claiming to offer a rebuttal
of modern evolutionary theory, only instead to engage in ‘bait and switch’
tactics by changing his target.
Returning again to the question of conflating
evolution as fact with evolution as theory, Harrison’s attempted dismissal of
“Darwinian evolution”, even if successful, does not mean the facts of common
descent and large-scale evolutionary change suddenly vanish, any more than
gravitational lensing, planetary motion, and frame dragging vanish if general
relativity was falsified. Even the intelligent design proponent and biochemist
Michael Behe, who denies the utility of the modern evolutionary synthesis,
nonetheless acknowledges the reality of common descent:
When two lineages share what appears to be an arbitrary genetic accident, the case for common descent becomes compelling, just as the case for plagiarism becomes overpowering when one writer makes the same unusual misspellings of another, within a copy of the same words. That sort of evidence is seen in the genomes of humans and chimpanzees. For examples, both humans and chimps have a broken copy of a gene that in other mammals helps make vitamin C As a result, neither humans nor chimps can make their own vitamin C. Of an ancestor of the two species originally sustained the mutation and then passed to both descendant species, that would neatly explain the situation…The same mistakes in the same gene in the same positions of both human and chimp DNA. If a common ancestor first sustained the mutational mistakes band subsequently gave rise to these two modern species, that would very readily account for both why both species have them how. It's hard to imagine how there could be stronger evidence for common ancestry of chimps and humans. (Emphasis mine) [14]
That
Harrison, a science educator, would
fail to see this point is particularly puzzling, and shows how adherence to a
dogma can blind a person to even the most obvious of facts. [15]
Harrison’s assertion that “Darwinian evolution…lacks the analytical
support enjoyed by the theories of true science.” is risible. If Harrison is referring to the fact of evolution, then there
has been no serious question in the scientific community about the reality of
common descent for over a century. Evolutionary biologist T.R. Gregory notes
that even in the 19th century, Darwin’s
list of evidence for the reality of an evolutionary history of life was
regarded as compelling, and continues:
Over the past 150 years, this initial list has been supplemented by countless observations in paleontology, comparative anatomy, developmental biology, molecular biology, and (most recently) comparative genomics, and through direct observations of evolutionary change in both natural and experimental populations. Each of thousands of peer-reviewed articles published every year in scientific journals provides further confirmation (though, as Futuyma…notes, “no biologist today would think of publishing a paper on ‘new evidence for evolution’ ... it simply hasn’t been an issue in scientific circles for more than a century”). Conversely, no reliable observation has ever been found to contradict the general notion of common descent. It should come as no surprise, then, that the scientific community at large has accepted evolutionary descent as a historical reality since Darwin’s time and considers it among the most reliably established and fundamentally important facts in all of science. (Emphasis mine) [16]
If he is referring to the modern evolutionary synthesis, which seeks to provide a theoretical explanation for the fact of evolution, then his assertion that it "lacks analytical support" betrays profound ignorance of modern evolutionary theory, which has considerable explanatory and predictive power.
Certainly, the existence of the highly successful (and mathematical!) discipline of population genetics, which lies at the heart of the modern synthetic theory alone is enough to dispose of Harrison's odd claim that evolution "lacks analytical support" [17] A classic example of the predictive power of evolutionary biology, one that has become readily apparent in the wake of the genomics revolution is the ability to test the hypothesis that if toothless mammals evolved from toothed mammals, we would expect to see decayed remnants of genes coding for tooth enamel. This is indeed the case:
Certainly, the existence of the highly successful (and mathematical!) discipline of population genetics, which lies at the heart of the modern synthetic theory alone is enough to dispose of Harrison's odd claim that evolution "lacks analytical support" [17] A classic example of the predictive power of evolutionary biology, one that has become readily apparent in the wake of the genomics revolution is the ability to test the hypothesis that if toothless mammals evolved from toothed mammals, we would expect to see decayed remnants of genes coding for tooth enamel. This is indeed the case:
Enamel is the hardest substance in the vertebrate body. One of the key proteins involved in enamel formation is enamelin. Most placental mammals have teeth that are capped with enamel, but there are also lineages without teeth (anteaters, pangolins, baleen whales) or with enamelless teeth (armadillos, sloths, aardvarks, pygmy and dwarf sperm whales). All toothless and enamelless mammals are descended from ancestral forms that possessed teeth with enamel. Given this ancestry, we predicted that mammalian species without teeth or with teeth that lack enamel would have copies of the gene that codes for the enamelin protein, but that the enamelin gene in these species would contain mutations that render it a nonfunctional pseudogene. To test this hypothesis, we sequenced most of the protein-coding region of the enamelin gene in all groups of placental mammals that lack teeth or have enamelless teeth. In every case, we discovered mutations in the enamelin gene that disrupt the proper reading frame that codes for the enamelin protein. Our results link evolutionary change at the molecular level to morphological change in the fossil record and also provide evidence for the enormous predictive power of Charles Darwin's theory of descent with modification. (Emphasis mine) [18]
His referral to "Darwinian evolution" betrays a lack of familiarity with modern evolutionary biology which has advanced considerably in the 150 years since Darwin first advanced his theory. As Gregory notes:
Modern evolutionary theory represents a multifaceted set of explanations for patterns observed both in contemporary populations and in deep time as revealed by the fossil record. Natural selection is considered by many to be the prime component of evolutionary theory and is the only workable mechanism ever proposed that is capable of accounting for the adaptive features of organisms. At the molecular level, nonadaptive mechanisms are recognized as highly significant, and there is also an increasing emphasis on changes due to processes such as genetic drift that differ from natural selection by being due to chance. [19]
Modern evolutionary theory is not the final word on the subject - no scientific theory by definition is complete and final, but it has considerable predictive and explanatory power. Harrison's statements about evolution are grossly inaccurate.
Abiogenesis and the creationist bait and switch
Abiogenesis and the creationist bait and switch
Harrison's declaration that "Darwinian evolution...lacks the analytical support enjoyed by true theories of science" as shown above is demonstrably false. The fact of command descent is attested by many independent lines of evidence from palaeontology to comparative genomics. The modern evolutionary synthesis has considerable explanatory and predictive power. His statement could be refuted by any competent biology undergraduate.
Having made this declaration, Harrison switches focus from evolutionary biology to abiogenesis, a classic bait and switch tactic of special creationists who trade on the general lack of scientific knowledge of fundamentalist laypeople who are unaware of this fact.
Having made this declaration, Harrison switches focus from evolutionary biology to abiogenesis, a classic bait and switch tactic of special creationists who trade on the general lack of scientific knowledge of fundamentalist laypeople who are unaware of this fact.
Apart from conflation of fact and theory and evolution with abiogenesis, the argument from personal incredulity, coupled with the argument from design, is one of the most frequently used special creationist fallacies. Typically, the special creationist will enumerate an aspect of the natural world, declare how it is 'too complex to have evolved', or proclaim that they cannot imagine how it evolved, then conclude that evolution is not true.
This approach ignores two main points:
This approach ignores two main points:
- Natural selection acting on random mutation is more than capable of effecting considerable design. As mentioned earlier, evolutionary algorithms are routinely used to evolve complex designs that can exceed that produced by an intelligent agent. Simulations of evolution using robots show how complex behaviour can evolve without conscious intervention. The design argument was laid to rest over 150 years ago. Special creationists have yet to catch up with this simple fact.
- The inability of one layperson to imagine a plausible evolutionary pathway for the evolution of the genetic code does not mean that evolution is false. Any layperson who declared that given his inability to reconcile quantum mechanics and general relativity, quantum gravity was impossible would not be taken seriously.
In fact, Harrison shows a profound lack of understanding of the realities of the evolution of the genetic code and the first cell:
Evolution wants us to believe that this code arose by chance at exactly the same time as the 1000+ proteins plus all the sugars, starches, fats and minerals that such a cell contains. Evolution believes that all these molecules randomly fell into place! It just happened. Even if you could find a free-living cell with only a tenth or a hundredth this size, the chance of exactly the right proteins, sugars, starches, fats and minerals plus an information molecule like DNA popping up by chance is hopelessly low. The critical point is that the information molecule had to arise simultaneously with all the cell’s molecules or it could not reproduce. Correlating a very large information molecule with the cell’s other molecules sounds like an intelligent act. Life is God’s intelligence in action.
Leaving aside the fact that what he is talking about here is abiogenesis, and not "Darwinian evolution". Harrison makes the mistake of thinking that origin of life researchers believe the first cell arose in one stage, and ignores the fact that the first proto-cell would be far less complex than even the simplest cells.

His comment betrays a profound misunderstanding of what origin of life researchers actually think. As molecular pharmacologist Ian Musgrage points out:
1) They calculate the probability of the formation of a "modern" protein, or even a complete bacterium with all "modern" proteins, by random events. This is not the abiogenesis theory at all.
2) They assume that there is a fixed number of proteins, with fixed sequences for each protein, that are required for life.
3) They calculate the probability of sequential trials, rather than simultaneous trials.
4) They misunderstand what is meant by a probability calculation.
5) They seriously underestimate the number of functional enzymes/ribozymes present in a group of random sequences. [20]
Harrison's failure to properly represent origin of life research seriously impacts on the credibility of his argument.
No scientist denies that abiogenesis is still an area in which many problems exist. There is no universally accepted theory for how the protocell was formed from organic molecules, though many hypotheses have been advanced. Some, such as the “RNA world” model [21] argue that nucleic acids formed first, while others postulate the formation of metabolic pathways first.
Stuart Kauffman’s work on autocatalytic sets or the Iron-Sulphur world are two prominent examples of this class of hypothesis. However, unlike evolutionary biology, we do not have a rigorous theory. Eugene Koonan summarises current research in abiogenesis in The Logic of Chance:
Stuart Kauffman’s work on autocatalytic sets or the Iron-Sulphur world are two prominent examples of this class of hypothesis. However, unlike evolutionary biology, we do not have a rigorous theory. Eugene Koonan summarises current research in abiogenesis in The Logic of Chance:
For all the effort, we do not currently have coherent and plausible models for the path from simple organic molecules to the first life forms. Most damningly, the powerful mechanisms of biological evolution were not available for all the stages preceding the emergence of replicator systems. Given all these major difficulties, it appears prudent to seriously consider radical alternatives for the origin of life. [22]
Present difficulties in a scientific discipline however do not mean that the problem is insoluble, as even a casual glance at the history of science would reveal. Dogmatism on the impossibility of abiogenesis ill-advised. For example. Lincoln and Joyce recently described the self-sustained replication of an RNA enzyme:
A long-standing research goal has been to devise a nonbiological system that undergoes replication in a self-sustained manner, brought about by enzymatic machinery that is part of the system being replicated. One way to realize this goal, inspired by the notion of primitive RNA-based life, would be for an RNA enzyme to catalyze the replication of RNA molecules, including the RNA enzyme itself. This has now been achieved in a cross-catalytic system involving two RNA enzymes that catalyze each other's synthesis from a total of four component substrates. [23]
Kamioka et al describe the creation of a structure capable of both replicating itself, and catalysing chemical reactions. This is significant for origin of life research, not only because they provide a starting point for creating complex molecules, but because these synthetic replicators have the possibility of mutation. [24]
Then there is the work of Hungarian biologist Tibor Gánti, whose 'chemotron model' provides a compelling model for the simplest system that can be called "living".

The chemoton (Ganti, 1984). The metabolic subsystem, with intermediates Ai, is an autocatalytic chemical cycle, consuming X as nutrient and producing Y as waste material; pVn is a polymer of n molecules of V', which undergoes template replication; R is a condensation byproduct of this replication, needed to turn r into T, the membranogenic molecule; the symbol Tm represents a bilayer membrane composed of m units made of T molecules. It can be shown that such a system can grow and divide spontaneously. (From John Maynard Smith and Eörs Szathmáry, The Major Transitions in Evolution (1995: OUP), p 21)
Gert Korthof, in a review of Gánti's book 'The Principles of Life' observes:
What is the simplest system that still can be called a living system? Plants and animals are composed of cells. Cells are the building blocks of life. So we need to focus on cells. But single cell organisms are still too complex. They can be simplified still further and still be called living. The entity that is stripped of all the unnecessary properties and is still alive is 'minimal life'. Gánti constructed an abstract model that captured minimal life. He called it the Chemoton model. It is composed of 3 subsystems:
1. Chemical motor system: A soft chemical self-reproducing system capable of synthesising chemical substances for itself as well as for the other two systems [metabolism. Example: proteins ]2. Chemical boundary system: a soft chemical system which is capable of spatial separation, of being selectively permeable to chemical substances, and of growth in the presence of its raw materials [ membrane. Example: lipids ]3. Chemical information system: a chemical system which is capable of self-reproduction in the presence of the appropriate raw materials [information. Example: RNA/DNA ]
Note: the 3 components superficially look like the three parts of the prokaryotic or eukaryotic cell: cytoplasm, membrane, nucleus. However, the chemoton model is not a model of the prokaryotic or eukaryotic cell, but a very general model for life; the simplest possible life.
The chemoton model does not contain enzymes (catalysts). It is a metabolism without enzymes. Since there are no enzymes, there is no need for the genetic code. This simplifies the chemoton model significantly. "Gánti liberated himself from the burden of the genetic code" says Szathmáry.
The chemoton model fulfils the 5 absolute life criteria. The chemoton is a unit, because deleting one of its subsystems reduces it to a chemical system. The chemical motor is equivalent with metabolism. The chemical motor is inherently stable (described in the book). The fourth criterion is fulfilled by the information carrying subsystem and the program control is present in the chemoton (described in the book). [25]
The point of citing Gánti’s chemotron model is not to argue that this is what the earliest living cell would have looked like, but to point out that origin of life researchers do not think that the first form of life was a complex cell, and did not form magically. Gánti’s chemotron model shows the level of simplicity that origin of life researchers postulate, and these are considerably more simple than obligate intracellular bacteria such as the Rickettsia, and show why Harrison's comparison with current living cells is invalid, and betrays his superficial grasp of the subject on which he pontificates.
This can be seen with the absence of any reference to experiments designed to shed light on abiogenesis such as Nobel Laureate Jack Szostak, who is one of the leading origin of life experts. Recently, Szostak announced the creation of a plausible prebiotic protocell in which nonenzymatic RNA copying could take place:
Efforts to recreate a prebiotically plausible protocell, in which RNA replication occurs within a fatty acid vesicle, have been stalled by the destabilizing effect of Mg2+ on fatty acid membranes. Here we report that the presence of citrate protects fatty acid membranes from the disruptive effects of high Mg2+ ion concentrations while allowing RNA copying to proceed, while also protecting single-stranded RNA from Mg2+-catalyzed degradation. This combination of properties has allowed us to demonstrate the chemical copying of RNA templates inside fatty acid vesicles, which in turn allows for an increase in copying efficiency by bathing the vesicles in a continuously refreshed solution of activated nucleotides. [26]
Again, the point of this is not to claim that abiogenesis has been solved. It has not, and the intellectually honest approach is to recognise that we do not know in detail how this occurred. Creationist misrepresentation of this problem however is anything but intellectually honest.
Evolution and Abiogenesis - Quote Mining
Harrison seeks to bolster his anti-evolution argument by appealing to the authority of prominent evolutionary biologists. Keep in mind that he has asserted that "Darwinian evolution...lacks the analytical support enjoyed by the theories of true science" but has so far resolutely failed to even properly differentiate between evolution as fact and evolution as theory, or differentiated between abiogenesis and evolution. He claims:
When asked to nominate the biggest questions facing evolution, three of ten leading biologists nominated the origin of life.
1 “The biggest gap in evolutionary theory remains the origin of life itself … the gap between … a collection of molecules and even the most primitive cell remains enormous ” (Chris Wills, UCSD).
2 Richard Dawkins admitted that the task is gargantuan . “Was the origin of life itself a probable event, and is life common in the universe?” Dawkins elsewhere explained that life may have been seeded on earth by intelligent aliens!
3 “The most profound unsolved problem in biology is the origin of life itself. We know a great deal about the creative chemistry of the early earth, but not yet enough to solve this problem” (Kenneth Miller, Brown University).
Harrison's failure to provide a reference for his quotes is shoddy research - he should provide a reference to allow readers to verify the provenance and accuracy of his quotation. It does not take much effort to find out that his source is a 2009 New Scientist article [27] in which several prominent evolutionary biologists were asked to point out what they thought were "the biggest gaps remaining in evolutionary theory."
Needless to say, none of these prominent evolutionary biologists believe that there is any doubt that evolution occurred, so it is highly misleading to cite them in such a way as to convey that impression.
Needless to say, none of these prominent evolutionary biologists believe that there is any doubt that evolution occurred, so it is highly misleading to cite them in such a way as to convey that impression.
It is worth noting that only three of the ten scientists quoted nominated abiogenesis as a major unsolved problem. In fact, when we look at some of the unsolved problems, it is painfully clear just how misleading Harrison's citation of this article is:
Frans de Waal
Why do humans blush? We're the only primate that does so in response to embarrassing situations (shame), or when caught in a lie (guilt), and one wonders why we needed such an obvious signal to communicate these self-conscious feelings. Blushing interferes with the unscrupulous manipulation of others. Were early humans subjected to selection pressures to keep them honest? What was its survival value?
Frans de Waal is Charles Howard Candler Professor of Primate Behavior at Emory University, Atlanta, Georgia
Richard Forty
We need to know more about the role of geographical isolation in stimulating the production of new species. Is this factor responsible for the greater part of biodiversity? How does evolution produce enormously species-rich genera, such as Eucalyptus, many of which can coexist happily in the same habitat? I hope that this anniversary will result in a new impetus for field-based biology and palaeontology to answer such questions.
Richard Fortey is in the department of palaeontology at the Natural History Museum in London
Chris Stringer
We still don't know what the last common ancestor of humans and chimpanzees looked like, where and how it lived, and what processes sent us down our separate evolutionary paths. There are now a few important fossils from Africa in the likely time period of between 7 and 5 million years ago, but for me they have not necessarily brought us close to an answer. More fossil discoveries – and perhaps research on the living species, including further genetic data – are needed.
Chris Stringer is in the department of palaeontology at the Natural History Museum, London [28]
Ignoring the fact that if only 3 of the 10 scientists thought abiogenesis was the biggest unsolved problem in evolution, the lack of a universally accepted theory of abiogenesis can hardly be seen to be an 'evolution killer', the context of these quotes shows that the unsolved problems are just that - problems for which no solution has yet been found. None of the scientists are declaring that evolution has been falsified by these problems, which is unfortunately the impression the layperson will infer from reading Harrison's article.
By far the most disturbing part of Harrison's citation of the New Scientist article is the selective way in which he quoted Kenneth Miller. Harrison's citation is in blue:
Kenneth Miller
I don't think there are basic “gaps” in the theory of evolution, which has proven to be a remarkably flexible scientific framework, brilliantly accommodating new data and even new fields of science, like molecular genetics. However, the most profound unsolved problem in biology is the origin of life itself. We know a great deal about the creative chemistry of the early Earth, but not yet enough to solve this problem.
Kenneth Miller is professor of biology at Brown University, Providence, Rhode Island
This is inexcusably dishonest. Harrison omits the first part of Miller's quotation in which he states that in his opinion there are no basic gaps in evolutionary theory, which Miller believes, contrary to Harrison's assertion that evolution "lacks the analytical support enjoyed by the theories of true science" is a "remarkably flexible scientific framework, brilliantly accommodating new data and even new fields of science".
Given that Miller is a respected cell biologist while Harrison is not even a scientist, it is hardly unreasonable to point out that Miller's opinion is considerably more informed and reliable than that provided by Harrison, whose quote mining, failure to cite references, bait and switch tactics, and conflation of evolution and abiogenesis reflect poorly on his commitment to intellectual honesty.
Given that Miller is a respected cell biologist while Harrison is not even a scientist, it is hardly unreasonable to point out that Miller's opinion is considerably more informed and reliable than that provided by Harrison, whose quote mining, failure to cite references, bait and switch tactics, and conflation of evolution and abiogenesis reflect poorly on his commitment to intellectual honesty.
Conclusion
Given that there has been no serious doubt in mainstream scientific circles about the reality of common descent and large-scale evolutionary change, the burden of proof lies solely on evolution denialists such as Harrison to make their case. From what we've seen in his February 2015 article in The Christadelphian, it is safe to conclude he has not met that burden.
References
- Bramhill A “Preface: ‘Creation – A sense of immediacy’ The Christadelphian (2015) 152
- Any assertion that humanity needs to be descended exclusively from Adam and Eve in order to genetically inherit the guilt and/or the consequences of Adam’s sin owes everything to the heresy of Original Sin, and nothing to a proper understanding of the Bible.
- Harrison A “The power of God’s wisdom in creation” The Christadelphian (2015) 152:65-68
- Critically, he is not a scientist, which means his anti-evolutionary arguments not only run counter to the overwhelming, long-standing scientific consensus on the reality of common descent and large-scale evolutionary change, but are made by an educated layperson with no in-depth understanding of the scientific disciplines which demonstrate the reality of common descent.
- Musgrave I “Lies, Damned Lies, Statistics, and Probability of Abiogenesis Calculations” TalkOrigins http://www.talkorigins.org/faqs/abioprob/abioprob.html
- Harrison, op cit p 65
- There are copious references to God sending the rain and snow, but the principles of atmospheric physics are well known, and allow us to understand how rain and snow form. God indeed sends the rain, but works through natural second causes. Furthermore in Ex 7:17-18, God states that he would strike the water with the staff in his hand in order to turn it to blood, yet God did not literally strike the Nile, but employed secondary methods (Aaron) to achieve his aims.
- Avise J.C. “Footprints of nonsentient design inside the human genome” Proc. Natl. Acad. Sci. USA (2010) 107(S2):8969-8976
- http://www.talkorigins.org/faqs/genalg/genalg.html
- Held L.I “Quirks of Human Anatomy. An Evo-Devo Look at the Human Body” (2009: Cambridge University Press)
- Schneider, T. D., 2000. Evolution of biological information. Nucleic Acids Research 28(14): 2794-2799
- Moran L "How do the IDiots Explain the Origin of Life?" Sandwalk October 23 2013
- Stephen Freeland “The Evolutionary Origins of Genetic Information, Part 3” BioLogos Blog August 5 2013
- Behe M The Edge of Evolution: The Search for the Limits of Darwinism (2007: Free Press) p 71-72
- It also raises significant questions about the quality of article review at The Christadelphian, when an article so profoundly flawed, misleading, and inaccurate makes it into print.
- Gregory T.R. “Evolution as Fact, Theory and Path” Evo Edu Outreach (2008) 1:46-52
- See for example Felsenstein J "Theoretical Evolutionary Genetics" for an example of the "analytical support" that the modern synthetic theory enjoys.
- Meredith, R.W., Gatesy, J., Murphy, W.J., Ryder, O.A., and Springer, M.S. 2009. Molecular Decay of the Tooth Gene Enamelin (ENAM) Mirrors the Loss of Enamel in the Fossil Record of Placental Mammals. PLoS Genetics 5(9): e1000634.
- Gregory op cit p 50
- Musgrave loc cit
- Cech T.R. The RNA Worlds in Context (2011) Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a006742
- Koonin E.V The Logic of Chance: The Nature and Origin of Biological Evolution (2011 FT Press)
- Lincoln T.A., Joyce G.R Self-Sustained Replication of an RNA Enzyme Science (2009) 323 (5918): 1229-1232
- Kamioka S et al “Autocatalysis and organocatalysis with synthetic structures” Proc. Natl. Acad. Sci. USA (2010) 541-544.
- Korthof G Review: "The Principles of Life" 29th Dec 2003 (updated 19th July 2013)
- Adamala K, Szostak JW Nonenzymatic Template-Directed RNA Synthesis Inside Model Protocells Science (2013) 342:1098-1100
- "Evolution's Final Frontier" New Scientist (2009) 201:41-43
- ibid, p 41-42

