Homo naledi - a spectacular new milestone in human evolution
We have another possible addition
to the human family - Homo naledi, a small-brained hominin whose skeleton is a
mix of ancient and modern details. The two astounding things about this latest
discovery are the sheer number of fossils found, and the fact that it
deliberately disposed of its dead, providing evidence of deliberate,
intelligent behaviour. The skeletal anatomy and evidence that it disposed of
its dead show that this is not just another ape, while its small cranial size
and mosaic anatomy show that it is definitely not Homo sapiens. The evidence
for human evolution was already compelling well before this, so this discovery
does not change the big picture. Rather, it serves to remind us why
fundamentalist opposition to human evolution is made despite, not because of
the evidence.
Two years after their discovery
of hominin remains in the Rising Star Cave system north of Johannesburg, South
Africa, Lee Berger and his colleagues presented their findings in two papers
[1-2] in the journal eLife. Irrespective of their classification, the sheer
volume of hominin fossils discovered, at over 1500, makes this one of the most
significant palaeoanthropological discoveries ever

While
primitive in some respects, the face, skull, and teeth show enough
modern features to justify H. naledi's placement in the genus Homo.
Artist Gurche spent some 700 hours reconstructing the head from bone
scans, using bear fur for hair.
Aside from these limited faunal materials, the Dinaledi collection is entirely composed of hominin skeletal and dental remains. The collection so far comprises 1550 fossil hominin specimens, this number includes 1413 bone specimens and 137 isolated dental specimens; an additional 53 teeth are present in mandibular or maxillary bone specimens. Aside from the fragmentary rodent teeth, all dental crowns (n = 179) are hominin, recovered both from surface collection and excavation. Likewise, aside from the few bird elements, all morphologically informative bone specimens are clearly hominin. In all cases where elements are repeated in the sample, they are morphologically homogeneous, with variation consistent with body size and sex differences within a single population. These remains represent a minimum of 15 hominin individuals, as indicated by the repetition and presence of deciduous and adult dental elements [3]

Figure 1. Dinaledi skeletal specimens.
The figure includes approximately all of the material incorporated in this diagnosis, including the holotype specimen, paratypes and referred material. These make up 737 partial or complete anatomical elements, many of which consist of several refitted specimens. Specimens not identified to element, such as non-diagnostic long bone or cranial fragments, and a subset of fragile specimens are not shown here. The ‘skeleton’ layout in the center of the photo is a composite of elements that represent multiple individuals. This view is foreshortened; the table upon which the bones are arranged is 120-cm wide for scale.
DOI: http://dx.doi.org/10.7554/eLife.09560.003
While its cranial capacity of 513 cc is well below that of ours at 1300 cc, H. naledi is described by its discoverers as having significant morphological differences both from the australopithecines and other members of genus Homo. Its hand anatomy according to Berger et al “possesses a combination of primitive and derived features not seen in the hand of any other hominin.” Similarly, its foot differs from members of both Australopithecus and Homo. The authors note that
H. naledi exhibits anatomical features shared with Australopithecus, other features shared with Homo, with several features not otherwise known in any hominin species. This anatomical mosaic is reflected in different regions of the skeleton. The morphology of the cranium, mandible, and dentition is mostly consistent with the genus Homo, but the brain size of H. naledi is within the range of Australopithecus. The lower limb is largely Homo-like, and the foot and ankle are particularly human in their configuration, but the pelvis appears to be flared markedly like that of Au. afarensis. The wrists, fingertips, and proportions of the fingers are shared mainly with Homo, but the proximal and intermediate manual phalanges are markedly curved, even to a greater degree than in any Australopithecus. The shoulders are configured largely like those of australopiths. The vertebrae are most similar to Pleistocene members of the genus Homo, whereas the ribcage is wide distally like Au. afarensis. [4]

Figure 2. Holotype specimen of Homo naledi, Dinaledi Hominin 1 (DH1).
U.W. 101-1473 cranium in (A) posterior and (B) frontal views (frontal view minus the frontal fragment to show calvaria interior). U.W. 101-1277 maxilla in (C) medial, (D) frontal, (E) superior, and (F) occlusal views. (G) U.W. 101-1473 cranium in anatomical alignment with occluded U.W. 101-1277 maxilla and U.W. 101-1261 mandible in left lateral view. U.W. 101-1277 mandible in (H) occlusal, (I) basal, (J) right lateral, and (K) anterior views. Scale bar = 10 cm.
DOI: http://dx.doi.org/10.7554/eLife.09560.019
While it is trivial to say that
every fossil is transitional, H. naledi, with its mosaic of features similar to
Homo and Australopithecius, as well as features unique to it is an excellent
case of a transitional hominin fossil, with a mix of primitive and advanced
features
This species combines a humanlike body size and stature with an australopith-sized brain; features of the shoulder and hand apparently well-suited for climbing with humanlike hand and wrist adaptations for manipulation; australopith-like hip mechanics with humanlike terrestrial adaptations of the foot and lower limb; small dentition with primitive dental proportions. In light of this evidence from complete skeletal samples, we must abandon the expectation that any small fragment of the anatomy can provide singular insight about the evolutionary relationships of fossil hominins. [5]
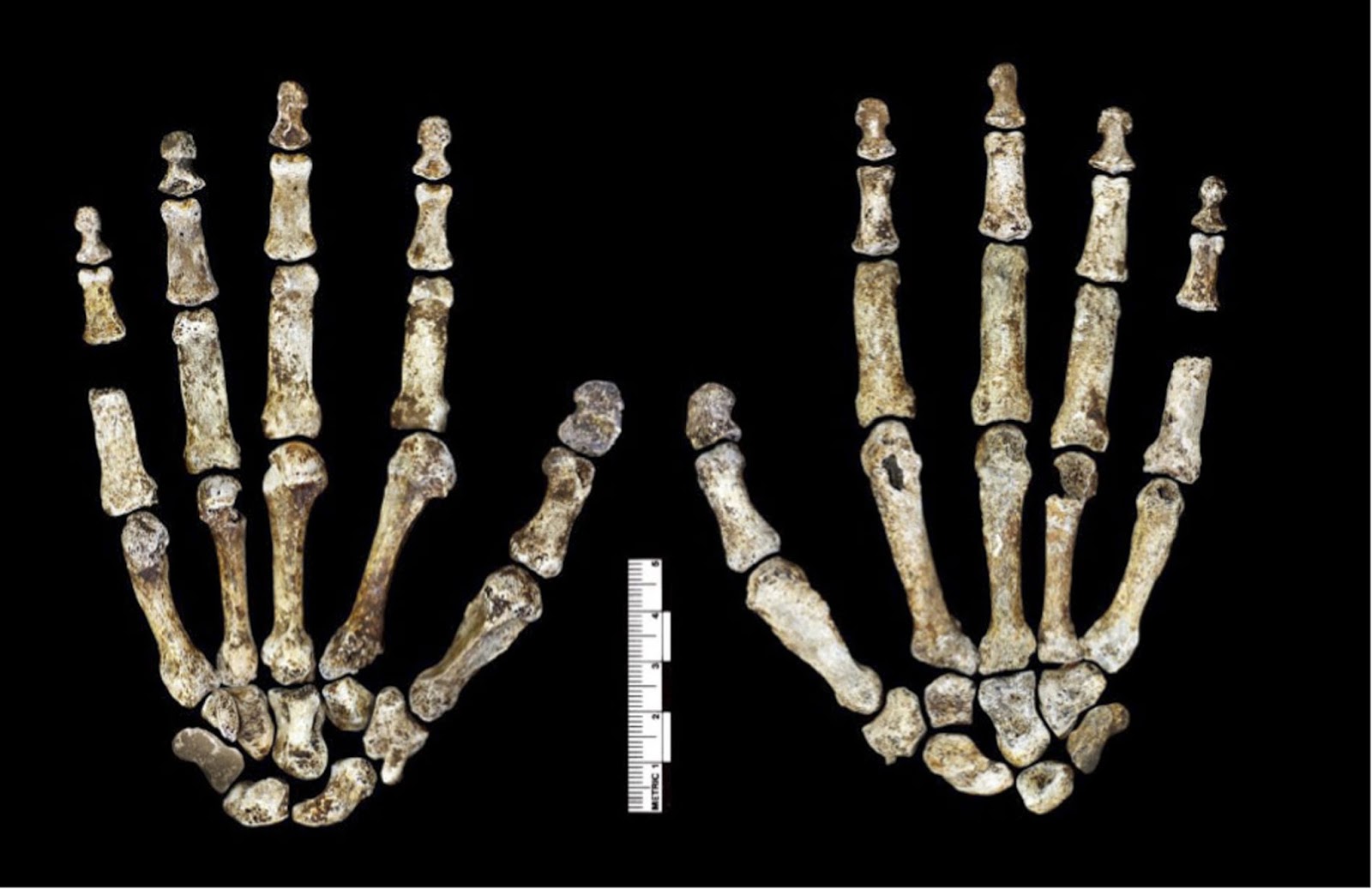
Figure 6. Hand 1.
Palmar view on left; dorsal view on right. This hand was discovered in articulation and all bones are represented except for the pisiform. The proportions of digits are humanlike and visually apparent, as are the expanded distal apical tufts on all digits, the robust pollical ray, and the unique first metacarpal morphology.
DOI: http://dx.doi.org/10.7554/eLife.09560.008
As yet, the discovery is undated,
which means that certain hypotheses about its significance for human evolution
cannot be fully tested until a secure date for the fossils is established. One
thing that appears to be fairly well established is that these fossils were
deliberately placed there. One would expect a mix of hominin and non-hominin
bones if they were washed into the cave system, yet as mentioned at the start,
apart from a tiny number of bones, the remains are entirely hominin.
The difficulty in accessing the
fossil remains also bears witness to the high probability that they were
deliberately placed there
The fossil-bearing chamber, named the Dinaledi Chamber (‘Chamber of Stars’ in the Sotho language; Figure 2A), is ~30 m below surface and ~80 m, in a straight line, away from the present, nearest entrance to the cave (Figure 2B). It is situated in the central part of the system and was found during speleological surveys (see methodology section). The only identified access point into the Dinaledi Chamber involves an exposed, ~15 m climb from the bottom of a large ante-chamber (the Dragon's Back Chamber), up the side of a sharp-edged dolomite block that has dislodged from the roof (the Dragon's Back; Figure 2B). From the top of the Dragon's Back, the Dinaledi Chamber is accessed via a narrow, northeast-oriented vertical fissure, and involves a ~12 m vertical climb down, with squeezes as tight as ~20 cm, to reach the floor (Figure 2B,C). The main passage forming the Dinaledi Chamber is ~25–50 cm wide at its narrowest and ~10 m long, and expands in width near the intersections with cross-cutting passages (Figure 2C). The roof of both the Dinaledi and Dragon's Back chambers is formed by the capping chert (Figure 2B). [6]

Figure 2. Geological map and cross-section of the Rising Star cave system.
(A) Geological Map showing the distribution of chert-free dolomite and fracture systems controlling the cave. Inset shows the location of the Cradle of Humankind in southern Africa; (B) Northeast-Southwest, schematic cross section through the cave system, relative to several chert marker horizons; (C) Detailed map of the Dinaledi Chamber showing the orientation of the floor and the position of the excavation and sampling sites.
DOI: http://dx.doi.org/10.7554/eLife.09561.004
(A) Geological Map showing the distribution of chert-free dolomite and fracture systems controlling the cave. Inset shows the location of the Cradle of Humankind in southern Africa; (B) Northeast-Southwest, schematic cross section through the cave system, relative to several chert marker horizons; (C) Detailed map of the Dinaledi Chamber showing the orientation of the floor and the position of the excavation and sampling sites.
DOI: http://dx.doi.org/10.7554/eLife.09561.004
Is it possible that there was a
more accessible path to the caves that has over time been blocked? The
researchers claim that there is no evidence for this:
An exhaustive search by a professional caving team and researchers has failed to find any other plausible access points into the Dinaledi Chamber, and there is no evidence to suggest that an older, now sealed, entrance to the chamber ever existed. Furthermore, detailed surface mapping of the landscape overlying the Rising Star cave system (Figure 2A) illustrates that no large flowstone-filled fractures occur in the region above the Dinaledi Chamber. [6]
That these bodies appear to have been deliberately placed in this cave (burial
is probably too strong a word) positively shouts a level of intelligence that
would set them apart from known ape behaviour, but their small skull and mix of
primitive and more modern anatomical features definitely brands them as not a
member of Homo sapiens. The implications of this for special creationism are
obvious. Furthermore, given that there
are still many more fossils in the Rising
Star Cave
system remaining uncollected, this is only the beginning chapter of this intriguing
story in human evolution, which has potentially significant implications, as the
respected palaeoanthropologist Chris Stringer in his commentary on the discovery
notes
Even without date information, the mosaic nature of the H. naledi skeletons provides yet another indication that the genus Homo had complex origins. The individual mix of primitive and derived characteristics in different fossils perhaps even indicates that the genus Homo might be ‘polyphyletic’: in other words, some members of the genus might have originated independently in different regions of Africa. If this is the case, it would mean that the species currently placed within the genus Homo would need to be reassessed (Dembo et al., 2015; Hublin, 2015). While many have concentrated on East Africa as the key and perhaps sole region for the origins of the genus Homo, the continuing surprises emerging from further south remind us that Africa is a huge continent that even now is largely unexplored for its early human fossils. [8]

Comparison of skull features of Homo naledi and other early human species.
Replica crania of (left to right) Homo habilis (KNM-ER 1813, Koobi Fora, Kenya ∼1.8 million years old), an early Homo erectus (D2700, Dmanisi, Georgia ∼1.8 million years old) and Homo floresiensis (Liang Bua 1, Indonesia ∼20,000 years old) are compared with actual fragments of cranial material of H. naledi that have been overlaid on a virtual reconstruction (far right; note some of the images of H. naledi material have been reversed). In each case, the crania are labelled with the typical features of each species. For example, while the adult brain volume of modern humans (Homo sapiens) is typically between 1000 and 1500 cubic centimetres (cc), H. habilis ranged from about 510 to >700 cc, H. erectus from about 550 to >1100 cc, H. floresiensis about 426 cc, and H. naledi between 466 and 560 cc. Furthermore, in modern humans, the occipital bone (at the back of the skull) is typically evenly rounded in profile, whereas in some early humans such as H. erectus, the upper and lower portions of the occipital are sharply angled to each other (i.e., ‘flexed’), and there is a strong ridge of bone running across the angulated region (called a transverse torus). Reference: eLife 2015;4:e10627
eLife 2015;4:e10627
Unsurprisingly, special
creationists have not let their lack of palaeoanthropological competence or the
fact they have not examined first-hand the fossils and published their findings
from offering the usual vacuous hand-waving dismissals. The YEC extremist
organisation Answers in Genesis has declared them “variations that developed
among apes”, a view that not only is flatly contradicted by the fossils which are
clearly hominin, but is also contradicted by Kurt Wise, a prominent YEC who is quoted
as saying:
“I think the case is very strong that these fossils are not just of the genus Homo, but are actually fully human (meaning they are descendants of Adam and Eve),” said Kurt Wise, director of the Center for Creation Research at Truett-McConnell College.
Fully human? Fully ape? When the
YECs can’t even agree among themselves on how to classify the same find, not
only does it show that they cannot be trusted to provide an authoritative,
informed comment on these discoveries, [9] it implicitly testifies to their
status as transitional fossils, given that the YEC denies a priori the concept
of transitional fossils, and is forced to shoehorn them into either a ‘human’
or an ‘ape’ category.
Given the special creationist
response to Homo naledi has been confused, superficial, and contradictory, one
hardly expects the anti-evolutionists in our community, who uncritically follow
the lead of these special creationist organisations, to provide any incisive,
informed commentary. One sadly expects it to be more like this comment from a Christadelphian science denialist Facebook page
Yeah, but they also profess that there were hominids too, even though they are ignoring Gen 2:7. So with the so called recent find in Africa, all someone needs to do is just take the remains of a human skull or gorilla skull, grab some plaster of paris, and whalla, a recently discovered species. Then those who "discovered" it can claim a meal ticket for as long as it lasts.
It hardly needs stressing that
the author of this comment is utterly uninformed on the subject, as the
researchers most certainly did not ‘take the remains of a human skull or
gorilla skull’ and coat it with plaster of paris. It is clear that he has not
read the original article, and is merely throwing uninformed speculation up in
order to fend off evidence that completely destroys his worldview. For the
record, here is a comparison of the Homo naledi skull and a human skull,
followed by a comparison of a modern human skull with a gorilla skull, which are enough to show the vacuity of the claims
expressed.

The braincase of this composite male skull of H. naledi measures a mere 560 cubic centimeters in volume—less than half that of the modern human skull behind it.
ART BY STEFAN FICHTEL. SOURCES: LEE BERGER AND PETER SCHMID, WITS; JOHN HAWKS, UNIVERSITY OF WISCONSIN-MADISON (Source: http://news.nationalgeographic.com/2015/09/150910-human-evolution-change/)
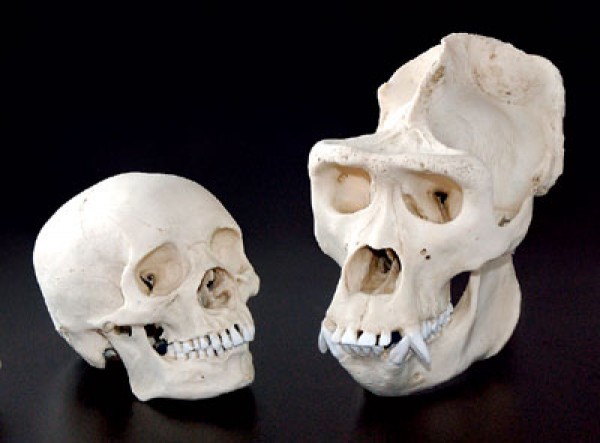 |
| Human and gorilla skull. Source |
Conclusion
While none of this changes the
basic fact of human evolution, depending on the date of these hominins, it
serves to show that the origin of the genus Homo was complex. Whether Homo
naledi is a genuine species, or is better classified as another member of the
genus is something that only further research will show. However, for those in
our community who continue to hide from the palaeoanthropolological evidence,
or peddle the long-rebutted claims of YEC organisations, this discovery should
serve as a wake-up call that such denial of reality is increasingly difficult. An
honest admission that we were wrong, coupled with an apology for persecuting and
slandering those who have been pointing out these facts, is long overdue.
References
1. Berger, Lee R. et al. "Homo naledi, a new species of
the genus Homo from the Dinaledi Chamber,
South Africa".
eLife (2015) 4.doi: http://dx.doi.org/10.7554/eLife.09560
2. Dirks Paul HGM. et al.
« Geological and taphonomic context for the new hominin species Homo
naledi from the Dinaledi Chamber,
South Africa”
eLife (2015) 4 doi: http://dx.doi.org/10.7554/eLife.09561
3. Berger, op cit.
4. ibid
5. ibid
6. Dirks, op cit
7. ibid
8. Stringer C “Human evolution: The
many mysteries of Homo naledi” eLife (2015) 4 doi: http://dx.doi.org/10.7554/eLife.10627
9. This is clearly indicated by
the absolute failure of special creationists to classify hominin fossils as ‘human’
or ‘ape’. See http://www.talkorigins.org/faqs/homs/compare.html

