Suboptimal design and the problem it poses for special creation
As a medical doctor, my profession brings me face to face with the stark fact that the human body, from the gross anatomical level down to the genomic, abounds with many features that either predispose us to suffering, disease, and even premature death. Reconciling this with special creation is frankly impossible for those who believe that every species - including humans - was individually hand-crafted. YEC arguments that suffering entered the world as a consequence of Adam's sin are immediately falsified by the overwhelming evidence of parasitism, predation, and disease hundreds of millions of years before the appearance of Homo sapiens (let alone Adam). Many people are unfamiliar with the considerable evidence for suboptimal design in the human body which directly or indirectly causes morbidity and mortality, so a post on this subject is definitely indicated, if only to educate those who conflate their uninspired, uninformed, fundamentalist reading of the Bible with the inspired word itself, creating a totally needless conflict between the overwhelming evidence for an ancient, evolving biosphere and an interpretation of the Bible which owes everything to biblicist ignorance and nothing to careful, informed study.
Light sensing cells in the vertebrate retina do not point towards the light, as one would righty imagine, but point away from the light. There are many problems arising from this design, the least of which is a blind spot in the retina to allow the blood vessels and nerve fibres which are in front of the light sensing cells to leave the eye. The human brain manages to edit out this deficit in our visual field so that we are normally unaware of it unless we specifically test for it. However, if the light sensing cells pointed towards the light, the blood vessels and nerves would be behind the retina, and the question of a blind spot would simply be a non-issue. Any special creationist claim that this is the only possible way to design an eye is blow out of the water by the cephalopod eye, which has a non-inverted retina, and therefore no blind spot.
 |
| Vertebrate and cephalopod eye. (Source) |
The existence of a blind spot is not the only consequence of an inverted retina. Unlike the cephalopod retina, the vertebrate retina is not firmly anchored to the back of the eyeball, which predisposes us to retinal detachment and visual impairment. This is not a trivial problem but rather is a genuine ophthalmological emergency as permanent visual loss or blindness results without surgical correction.
As the blood vessels are in front of the light sensing cells, any retinal haemorrhage will result in visual impairment. Another consequence of having blood vessels before the light sensing cells is the vision-threatening problem of diabetic retinopathy, caused by an increase in retinal blood vessel growth as a direct response to chronic lack of oxygen. As these blood vessels proliferate, they reduce visual acuity, and if not treated medically, can cause blindness.
Two other causes of visual impairment are macular oedema and macular degeneration, and these are directly attributable to a workaround required to produce an area of high-resolution vision to compensate for the inverted retina design. Neurologist Steven Novella notes:
"The macula is that part of the retina that has the densest concentration of rods and cones for detailed vision. Within the macula is a smaller area called the fovea which contains only cones and has the highest density of these receptors. The very existence of the macula, however, is a partial fix for the “backward” arrangement of retinal layers with the nerve and blood vessels between the receptors and the direction of light. This limits the density of rods and cones, and so the partial fix is to have one small area cleared of nerves and blood vessels where rods and cones can be denser. However, if the human retina were designed like that of the squid and other cephalopods, this would not be necessary.
"The dependence of the human eye on the macular for sharp vision creates a vulnerability, for any problem with that small area will have a dramatic effect on visual acuity. The rest of the retina will not be able to adequately compensate for the loss or compromise of the macula because the density of rods and cones is just too diffuse.
"Macular degeneration is the most common cause of blindness. Although the cause is often unknown, its severe effects on vision are a consequence of the need for a macula as a partial fix for the poor retinal design." [1]
 |
| Given that the macula contains the fovea, the site of high resolution vision, damage to one tiny part of the retina can result in marked visual impairment. (Source: Wikipedia) |
Far from being a minor problem, the inverted human retina is an example of suboptimal design that directly contributes to visual impairment and blindness. The existence of the objectively superior cephalopod retina not only shows that a better way of designing eyes exists, but also mocks the YEC argument that the suboptimal design is a consequence of an Edenic curse. Why were vertebrates cursed with a flawed retinal design, while the cephalopod retina escaped unscathed?
Problems with plumbing
A large number of medical problems arise from anatomical quirks that predispose humans to blockages in the urinary and gastrointestinal tracts, problems that could have been easily avoided if these quirks did not exist. For example, the human pancreas forms from two buds that fuse during embryogenesis. Occasionally, these two buds fuse around the duodenum, the first part of the small intestine, creating an annular pancreas. This can cause small bowel obstruction, and could easily have been avoided if the pancreas formed from a single bud.
Despite the fact that humans do not lay eggs, and therefore do not need a yolk sac, this structure nonetheless forms during pregnancy, being an inheritance from our reptile-like ancestors. (Further evidence of our egg-laying ancestry comes from the presence of the vitellogenin pseudogene [2], the decayed remnants of a gene that in egg-laying animals is involved in the production of egg yolk.)
The yolk sac is linked to the foetal small intestine via the omphalomesenteric duct which usually vanishes during the first trimester. In around 2% of the population, a remnant of the omphalomesenteric duct at its small intestine end persists as a Meckel's diverticulum. Complications of a Meckel's diverticulum include bleeding, obstruction, herniation, discharging umbilical sinus, and malignancy. Given that we do not need a yolk sac, the existence both of a structure that is not needed, as well as one that directly leads to complications in a significant minority of the population is impossible to reconcile with intelligent design.
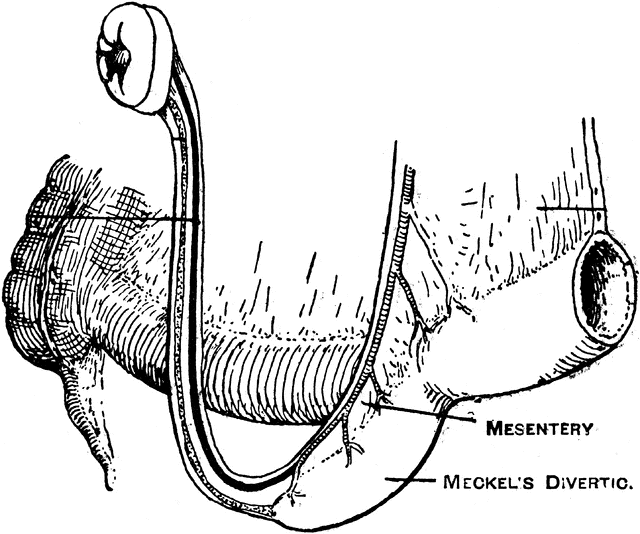 |
| Source |
The human prostate encircles the urethra, which guarantees that when the prostate enlarges (as it does in most men as they age), the urethra is compressed, resulting in impaired urinary flow. Occasionally, complete urinary retention results, which results not just in considerable pain and discomfort, but renal failure. This problem could be easily solved by connecting the prostate laterally to the urethra, preventing urinary obstruction.
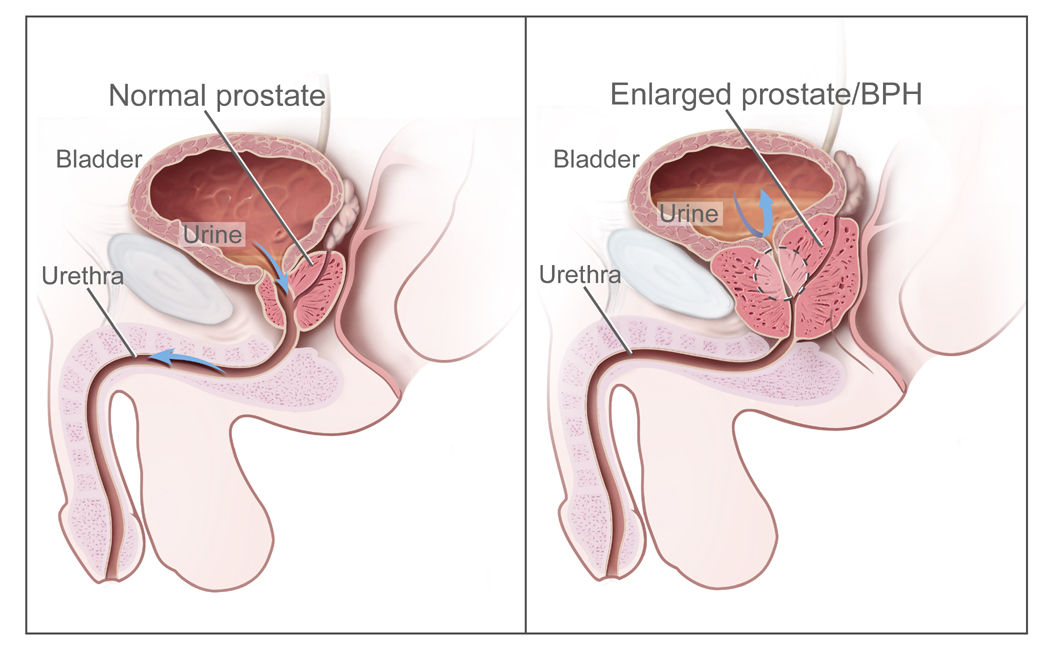 |
| Routing the urethra through the prostate means urinary obstruction is a real possibility with prostatic hypertrophy. (Source: Wikipedia) |
Another urogenital problem arises from the placement of the testicles outside the body. The testicles are actually originally inside the body, and during embryogenesis descend and exit the body. Apart from having sensitive structures located in a vulnerable position, their passage outside the body creates a weakness in the abdominal wall leading to a high predisposition for inguinal hernias, which if they strangulate can cause death if not surgically treated. In some male infants, the testes fail to descend properly, resulting in the phenomenon of cryptorchidism, where a testicle fails to descend into the scrotum. This is hardly trivial, as it results in an increased risk of infertility and testicular cancer. The testicles are outside the body as spermatogenesis does not occur at body heat. For an intelligent designer, the problem is simple - engineer spermatogenesis so that it can take place at body heat, and leave the testicles inside the body cavity. Many animals have internal testicles, so the presence of external testicles in humans (and other mammals) is again impossible to reconcile with intelligent design.
 |
| Source: Wikipedia |
This hardly exhausts the list of anatomical quirks that both lead to morbidity and mortality, and are readily corrected. Other examples include:
- Back pain arising from the co-option of a spine suited for quadrupeds for use by a biped, one that is also susceptible to disc prolapse.
- Ectopic pregnancy arising from a fertilised egg adhering to the fallopian tube. Direct connection of the ovaries to the uterus to allow direct release of the egg into the uterus for fertilisation would solve this life-threatening problem
- We are prone to sinus blockage and infection due to the way in which the sinuses drain which makes sense for a quadruped, but no sense for a biped.
- Unnecessarily long pathway of the recurrent laryngeal nerve, which bypasses the larynx, goes down into the chest, under the aorta, then back to the neck. This quirk is inherit by all mammals, and at its extreme (giraffe) results in several wasted metres of nerve. [3]
Suboptimal genomic design
These problems are not retracted to gross anatomy. They can be found at the genomic level. The most celebrated example is the fact that unlike many animals, we cannot synthesise our own vitamin C. This inborn error of metabolism means we need to find a reliable source of dietary vitamin C; if we do not get enough vitamin C, we will contract the potentially lethal disorder known as scurvy. Vitamin C is synthesised through the action of a number of enzymes on pre-existing chemicals. We have all the enzymes except for the last one: GULO, the gene that codes for L-gulonolactone oxidase, the final enzyme in the biosynthetic pathway is a pseudogene.
Other animals such as apes, monkeys, some fish, and guinea pigs also have a crippled L-gulonolactone oxidase gene. However, humans, apes, and monkeys have exactly the same error in GULO, a fact consistent with the pseudogenisation of GULO in the common ancestor of humans, apes, and monkeys. Special creationists have many problems to answer just on this point:
- Why were humans created with a biosynthetic pathway for vitamin C, an essential nutrient, that was broken?
- Why are other animals created with a functioning GULO gene?
- Why are humans, apes, and monkeys created with a GULO gene broken in exactly the same way, and with exactly the right pattern of point mutations that one would expect from a model of common descent?
Evolutionary biologist John Avise, in a recent PNAS paper has shown that genomic suboptimal design can be found right through to the very architecture of the genome, and the method by which genetic data is turned into proteins.
Introns, Exons, and splicing-related errors
Contrary to what many laypeople would imagine, most of the gene does not consist of protein-coding genetic material. In fact, genes consist of small protein-coding sections (exons) that are interleaved with much larger non-coding sections known as introns. When the cell needs to make a protein, the DNA copy is transcribed to an RNA copy. The intronic sections are removed, with the remaining exonic RNA spliced together, where, if it is a protein-coding gene, will be used as a template to make a protein at the ribosome. This extra genetic overhead comes at a significant metabolic cost of transcribing DNA that will eventually be snipped out and not used to directly code for a protein. While the existence of separate exons allows for the creation of some genetic novelty, this is far outweighed by the many genetic errors that are directly attributed to this process. Avise notes:
Nevertheless, for the sake of argument, let us assume that the metabolic costs imposed by introns are negligible. Do introns otherwise provide evidence of optimal genomic design? No, because premRNA processing also has opened vast opportunities for cellular mishaps in protein production. Such mishaps are not merely hypothetical. An astonishing discovery is that a large fraction (perhaps one-third) of all known human genetic disorders is attributable in at least some clinical cases to mutational blunders in how premRNA molecules are processed. For example, it has long been known that mutations at intron-exon borders often disrupt premRNA splicing in ways that alter gene products and lead to countless genetic disabilities, including various cancers and other metabolic defects. There is also good evidence that the number of introns in human genes is positively correlated with a gene’s probability of being a disease-causing agent. Avise summarizes many of the human genetic afflictions that have been documented (in particular clinical instances) to molecular errors in mRNA splicing at specifiable loci. These range from a variety of neurodegenerative diseases to debilitations of the circulatory, excretory, and other body systems. Many of these genetic disorders begin in infancy or early childhood; others are deferred to the elderly. The devastating symptoms of many such disorders, such as Lou Gehrig disease (amyotrophic lateral sclerosis), are simply horrible by any human standard. [4]
By eliminating introns, not only would one have enough space (and then some) to directly code for proteins produced by alternative splicing, one would eliminate the problems of genetic disease directly related to intron-expon border mutation.
Mitochondrial DNA and genetic disease
The main location of energy production in the cell is the mitochondrion, which is unique in being the only structure outside of the nucleus to have its own DNA (mtDNA). One would imagine that if the mitochondria have been given their own DNA, then the mtDNA would code for all the proteins involved in energy metabolism. This however is not the case:
A remarkable fact is that four of these five enzyme complexes are composed of combinations of polypeptides from the mitochondrial and nuclear genomes. In complex IV, for example, 3 of the 13 polypeptides are encoded by mitochondrial loci (COI, COII, and COIII), whereas the remaining polypeptides are encoded by nuclear genes. Only in complex II are all the necessary enzymatic subunits (four in this case) encoded by just one genome (the nuclear). Nuclear genes are also intimately involved in other basic mitochondrial functions. Indeed, mtDNA does not encode any of the proteins that are directly involved in its own replication, transcription, translation, surveillance, or repair. In short, mtDNA is just a tiny snippet of DNA that by itself would be absolutely helpless to itself and to the organism in which it is housed. [5]
Furthermore, the genetic code in mtDNA is subtly different from nuclear DNA. Furthermore, mtDNA endures up to ten times more mutations in a time interval than nuclear DNA. None of this makes any sense from the point of view of intelligent design, but is perfectly explicable from an evolutionary biology point of view, as Avise notes:
None of this makes any biological sense, except in the light of evolutionary science (which has discovered that modern mitochondria are remnants of a microbe that invaded or was engulfed by a protoeukaryotic cell in an endosymbiotic merger that took place billions of years ago). [6]
The utility of evolutionary biology to medicine cannot be overestimated, as an understanding of the evolutionary origin of the human body allows us to understand why it fails. Avise correctly notes that "an emerging paradigm is that many of the degenerative diseases of aging have their etiologies in mitochondria, either as deleterious mutations in the mtDNA molecules themselves or as operational flaws in nuclear-mitochondrial interactions." Understanding how and why such genetic diseases occur allows one to work out how to ameliorate the suffering arising from them, but also raises the very real problem of why, if the human genome was intelligently designed, is so much subioptimal design hard-wired into it. Avise again:
"The serious health problems that arise from mtDNA mutations immediately challenge any claim for omnipotent perfection in mitochondrial design. Perhaps these mutational aberrations can be viewed as unfortunate but inevitable byproducts of molecular complexity. However, the intellectual challenges for ID go much deeper. Considering the critical role of cellular energy production in human health and metabolic operations, why would an intelligent designer entrust so much of the production process to a mitochondrion, given the outrageous molecular features this organelle possesses? Why would a wise designer have imbued mtDNA with some but not all of the genes necessary to carry out its metabolic role (and then put the remaining genes in the nucleus instead)? Why would a wise engineer have put any crucial genes in a caustic cytoplasmic environment in which they are exposed routinely to high concentrations of mutagenic oxygen radicals? Why would he have dictated that the mitochondrial genetic code must differ from the nuclear genetic code, thereby precluding cross-translation between two genomes for which effective communication would seem to be highly desirable? Why would an intelligent designer have engineered mtDNA structures (e.g., closed-circular genome, no introns, no junk DNA, lack of binding histones) and mtDNA operations (e.g., little or no genetic recombination, production of a polygenic transcript, limited ability to mend itself, no self-sufficiency in transcription or translation) to differ so fundamentally from their counterpart features in the nuclear genome? In a nutshell, the underlying design of the whole mitochondrial operation seems to make no (theo)logical sense. Not only is the overall design of mtDNA suboptimal, but it appears downright ludicrous!" [7]
Avise is correct. Many genetic diseases directly arise from mitochondrial genetics, and if what we see is the result of intelligent design, it poses major problems for theology.
Jumping Genes and genetic disease
The metaphor of human genome as encyclopaedia is commonly advanced, but to be honest, it is an unhelpful one, given that the genes consist mostly of non-coding DNA which is spliced out during the process of creating a RNA copy from which a protein can be made. The encyclopaedia would actually consist of page after page of gibberish, interleaved by small snippets of useful text. In fact, most of the human genome consists of non-coding DNA, with at least two thirds of the human genome junk that is not essential for life. Around 45% of the human genome consists of mobile genetic elements called retrotransposons, that do nothing but copy and paste themselves randomly through the genome. As one can imagine, random pasting of genetic material into the genome is not good idea, as the potential exists for writing over essential genes, causing genetic disorder:
Mobile elements have the potential to cause human diseases by several mechanisms. When a mobile element inserts into a host genome, it normally does so at random with respect to whether or not its impact at the landing site will harm the host. If it happens to land in an exon, it can disrupt the reading frame of a functional gene with disastrous consequences. If it jumps into an intron or an intron-exon boundary, it may cause problems by altering how a gene product is spliced during RNA processing. If it inserts into a gene’s regulatory region, it can also cause serious mischief. The potential for harm by such insertional mutagenesis is great. It has been estimated, for example, that an L1 or Alu mobile element newly inserts somewhere in the genome in about 1–2% and 5%, respectively, of human births. Another problem is that when a mobile element lands in a functional gene, genetic instabilities are sometimes observed that result in deleted portions of the recipient locus. Several genetic disorders have been traced to genomic deletions associated with de novo insertions of mobile elements. Finally, mobile elements (or their immobile descendents that previously accumulated in the human genome) can also cause genomic disruptions via nonallelic homologous recombination. Serious metabolic disorders can result.
Despite the relatively recent discovery of mobile elements, the list of genetic disorders associated wholly or in part with their activities already is long. Still, any such list provides only a minimum estimate of these elements’ collective toll on human health. This is because some of the most serious medical difficulties probably arise so early in ontogeny as to cause miscarriages that normally will remain of unknown etiology. Indeed, most mobile elements are especially active in the germ line; thus, many of their deleterious effects probably register in gametic deaths and lowered fertility. [8]
Any minor benefit that arises from the emergence of genetic novelty from co-option of retrotransposon elements (and this benefit presupposes evolution, so it is not available for the special creationist to use) is far outweighed by the fact that inside the human genome are parasitic elements which can copy and paste themselves around the genome, causing disease in the process. Harmonising this with intelligent design is difficult at best.
Conclusion
The human body, from the gross anatomical level down to the genome is replete with suboptimal design that not only contributes to morbidity and mortality, but could have easily been avoided. None of this poses any problem for evolutionary creationism, as this is exactly what one would expect if evolution was the mechanism by which the diversity of life appeared. For the special creationist, such suboptimal design is a problem that simply cannot be explained away. The problem of suffering skewers special creationism, and only an evolutionary theodicy can solve the problem, as Avise (and others) have wisely noted:
Evolution by natural causes in effect emancipates religion from the shackles of theodicy. No longer need we agonize about why a Creator God is the world’s leading abortionist and mass murderer. No longer need we query a Creator God’s motives for debilitating countless innocents with horrific genetic conditions. No longer must we anguish about the interventionist motives of a supreme intelligence that permits gross evil and suffering in the world. No longer need we be tempted to blaspheme an omnipotent Deity by charging Him directly responsible for human frailties and physical shortcomings (including those that we now understand to be commonplace at molecular and biochemical levels). No longer need we blame a Creator God’s direct hand for any of these disturbing empirical facts. Instead, we can put the blame squarely on the agency of insentient natural evolutionary causation. From this perspective, the evolutionary sciences can become a welcome partner (rather than the conventionally perceived adversary) of mainstream religion. [9]
Exactly. The Christian who argues that the suboptimal design we see in nature was specially created is creating grounds for unbelief, not just on grounds of the exquisite suffering arising, but from the fact that such suboptimal design not just is consistent with common descent, but in many cases (particularly at the genomic) is inexplicable other than from common descent, a problem in that it means the world was created to look like it evolved. As one person once said, God is not the author of a lie, not even a white one. The Christian who accepts the witness of the natural world as a reliable guide to its formation will have no such problems, as she will recognise that just as God has granted free will to humanity, with the potential for good and evil, so has He granted the natural world freedom to evolve, with the same potential for exquisite and suboptimal design. [10]
References
1. Novella S "Suboptimal Optics: Vision Problems as Scars of Evolutionary History" Evo Edu Outreach (2008) 1:493–497
2. Brawand D, Wali W, and Kaessmann H, “Loss of Egg Yolk Genes in Mammals and the Origin of Lactation and Placentation,” PLoS Biology (2006) 6:0507–17.
3. Held L.I., Quirks of Human Anatomy: An Evo-Devo Look at the Human Body" (2009: Cambridge University Press)
4. Avise J.C., "Footprints of nonsentient design inside the human genome" Proc. Natl. Acad. Sci. USA (2010) 107:8969-8976
5. ibid., p 8974
6. ibid., p 8974
7. ibid., p 8974
8. ibid., p 8975
9. ibid, p 8975
10. Evolutionary biologist and former Catholic seminarian Francisco Ayala puts it well: "It was an unexpected turn of events for me, coming from conservative Spain, to discover that there was in the United States a strong creationist current that saw Darwin and the theory of evolution as contrary to religious beliefs. In Salamanca, in my theological studies, evolution had been perceived as a friend, not an enemy, of the Christian faith. Evolution, I learned in my theology classes, had provided theologians with the “missing link” in the explanation of evil in the world or, in theological parlance, evolution had solved the “theodicy” problem. A dictionary definition of theodicy is “defense of God’s goodness and omnipotence in view of the existence of evil.” See Ayala F "Darwin's Gift to Science and Religion: (2007: Joseph Henry Press)
This wisdom must be contrasted against the depressing example of the Christadelphian who combines utter scientific ignorance with a superficial fundamentalist reading of the text, along with a complete lack of insight into their many scientific, theological, and exegetical limitations. By doing so, they are idolising their own ignorance, and creating multiple stumbling blocks for the next generation who are told that YEC and Biblicism must be normative for all Christadelphians. When they discover that YEC is theological and scientific nonsense, too often they leave the faith. A large part of the blame for this lies fairly with the arrogance and ignorance of the average YEC proponent. James 3:1 is particularly pertinent here.


{kind=link}
Comments
Post a Comment