A criticism of Stephen Palmer's talks at the Coventry Creation Day - 12
Palaeoanthropology – more than just a few fossilised teeth
As with any attack on the fossil
record for evolution, the special creationist attack on palaeoanthropology
fails to comprehend what palaeontologists mean by transitional fossils.
Arguments referring to or implying the lack of ‘missing links’ betray
completely flawed ‘ladder view’ of evolution with bacteria at the bottom,
humans at the top, with millions of ‘links’ connecting bacteria to human in an
evolutionary ‘chain of progress’. This view is completely inaccurate, failing
as it does to recognise that evolution is not a ladder, but a tree, and that
what evolutionary biologists are looking for are not ‘missing links’ but
fossils that allow them to infer the underlying pattern of descent with
modification by virtue of the transitional features that they display. As
Louise Mead reminds us:
The concept of a “missing link” is an “archaic expression” tracing back to the Great Chain of Being, a view of the physical and metaphysical world as an unbroken chain. It was later temporalized by the evolutionary thought of the eighteenth and nineteenth century to the idea of evolution as a progressive climb up a ladder. These views of evolution create the false expectation that there should be fossil evidence showing “a complete chain of life from simple to complex. Creationists rely on such views to support their arguments against macroevolution, in particular by pointing out the “conspicuous” absence of “large numbers of intermediate fossil organisms”, using what is still unknown to question whether evolution has occurred.[1]
The other error special creationist make when attacking the fossil record is
their reference to ‘intermediate forms’ which again betrays his erroneous
ladder thinking. Meade continues:
Expectations of “intermediate” forms reflect neither Darwin’s original thoughts nor current thinking and practices in evolutionary biology and paleontology. Darwin (1859) was very clear on this. The ancestor of two living forms is unlikely to be found alive because it would have been outcompeted in most cases by newly adapting forms, and an extinct ancestor of two living forms would not be expected to look intermediate between them. Today, evolutionary biologists and paleontologists do not focus on finding “intermediates” but rather on reconstructing evolutionary relationships and history using shared derived characters, or synapomorphies. Willi Hennig revolutionized systematics in the 1960s with the introduction of cladistics, which ushered in a new method of phylogenetic analysis and a new approach to systematics. Instead of relying on a Linnaean system of classification, cladistics placed the focus on evolutionary history, specifically identifying features as ancestral (general) or derived (evolved after the lineage split from the ancestor). If a shared derived character, or synapomorphy, is found in two or more related organisms, it is inferred to have been present in their common ancestor, irrespective of whether or not there is a fossil record for that ancestor (Hennig 1966). Rather than trying to find the actual fossil corresponding to the “missing link” between lobe-fins and tetrapods, paleontologists instead look for fossils with characters or features important for an adaptive transition from life in an aquatic environment to life on land and that are shared as the result of common ancestry.[2] (Emphasis mine)
Therefore, palaeoanthropologists
are not looking for half-ape / half-human fossils, but examining the fossils
they find for evidence consistent with development of obligate bipedality,
reduced jaw size, increased cranial capacity, evidence of hands able to
exercise precision grip, and from analysing the data, they are able to
construct provisional evolutionary family trees for the human race, which can
be revised and refined as more fossil data emerges.
A commonly-shared image highlights the gross error made by special creationists:


As one can see, we do have
fossils that are close to the chimpanzee-human divergence which allow us to
infer a great deal about the human-ape common ancestor. Such special creationist
criticism of palaeoanthropology is not even wrong, because it fails to grasp
the essential point that evolution is not a ladder, but a tree. It also fails
to grasp the fact that the quality and quantity of human fossils discovered is
definitely more substantial than a few chipped teeth:
Opponents of scientific biology are fond of dismissing that record as a pathetic handful of controversial fragments. If that were so, this book would be a lot shorter. An often-repeated creationist canard insists that all known human fossils would fit on a billiard table. This was probably true in the 19th century, but it has not been true for a hundred years. Known human fossils number in the thousands and represent the remains of hundreds of individuals...Having seen most of the major collections of human fossils in the world's museums, we can assure our readers that those collections can no longer be laid out on a billiard table. It would be hard to cram them into a boxcar.[3]
Fossils near the base of the
human evolutionary tree
Currently, we have three fossils close to the human-chimpanzee split:
Currently, we have three fossils close to the human-chimpanzee split:
- Orrorin tugenensis
- Sahelanthropus tchadensis
- Ardipithecus ramidus
It is unlikely that any of these
species represents the common ancestor of humans and chimpanzees, but as I said
earlier, palaeontologists are more concerned with establishing relationships
between fossils in order to recreate the evolutionary tree. We can infer much
about the common ancestor even without the actual fossil, and the existence of
these fossils (and others) alone is enough to refute the special creationist
claims that there is no fossil evidence for human evolution.[4]
Orrorin tugenensis
Orrorin tugenensis was a
Miocene hominin found in Kenya in 2000 by Brigitte Senut and Martin Pickford[5] that has
been dated to approximately 6 million years ago. The remains discovered are
shown below, followed by a description from the paper:

Hominid
with jugal teeth smaller than those of Australopithecines; upper central
incisor large and not shovel-shaped, with thick enamel; upper canine short with
a shallow and narrow vertical mesial groove, apical height low; small
triangular upper M3s; corpus mandibularis relatively deep below M3; lower P4
with offset roots and oblique crown; small Homolike rectangular lower M2 and
M3s; thick enamel on lower cheek teeth; buccal notch well developed which
imparts a bilobate profile to the buccal surface; no cingulum on molars. Femur
with a spherical head rotated anteriorly, neck elongated and oval in section,
lesser trochanter medially salient with strong muscle insertions, deep digital
fossa; humerus with a vertical brachioradialis crest; proximal manual phalanx
curved; dentition small relative to body size.[6]
What that means is that they
found teeth, jaw fragments, a femur and a humerus. We have no skull sadly, but
we do have post-cranial data (bones other than the skull) which is important
for us to work out whether Orrorin tugenensis was bipedal. By examining
the thigh bone, we can work this out.

Morphological
comparisons among femora of or attributed to (A) P. troglodytes,
(B) O. tugenensis (BAR 1002'00), (C and D)Paranthropus
robustus (SK 97 and SK 82, reversed), (E) A. afarensis (A.L.
288-1ap), (F) Paranthropus boisei (KNM-ER 1503,
reversed), (G) early Homo (KNM-ER 1481), and (H)
modern H. sapiens. Like other early hominin femora (C to F), BAR
1002'00 (B) is distinct from those of modern humans (H) and great apes (A) in
having a long, anteroposteriorly narrow neck and wide proximal shaft.
Early Homo femora (G) have larger heads and broader necks
compared to early hominins. In addition to these features, modern human femora
(H) have short necks and mediolaterally narrow shafts. Scale bar, 2 cm.
Source: Richmond B.G., Jungers W.L. "Orrorin tugenensis Femoral Morphology and the Evolution of Hominin Bipedalism" Science (2008) 319:1662-1665
The authors of this paper felt that Orrorin tugenensis was bipedal, as they announced:
The postcranial evidence suggests that Orrorin tugenensis was already adapted to habitual or perhaps even obligate bipedalism when on the ground, but that it was also a good climber.[7]
The authors published a family tree, showing where they felt
Orrorin tugenensis was positioned:

Note that they place Orrorin tugenensis after the Australopithecine and Homo lines had diverged, close to this split.
Not everyone was convinced by
this. However, further studies performed on the bones strengthened the case for
those who believed Orrorin tugenensis was bipedal.
Our results show that the internal distribution of cortical bone in its femoral neck constitutes direct evidence for frequent bipedal posture and locomotion in this Late Miocene ancestor. In known features, external and internal, BAR100200 exhibits a total morphological pattern distinct from African apes, diagnostic of bipedal locomotion, and appropriate for a population standing at the dawn of the human lineage.[8]
Did that clinch the case? Not
quite. The CT scanning performed on the bones did not have very good resolution
- a recent study (2008) has clarified the issue, and concluded that Orrorin
tugenensis was quite likely bipedal, but was not more closely related to Homo
than to Australopithecus:
In light of the marked changes in femoral anatomy from at least 2 Ma to the present, the close morphological similarity between femora of O. tugenensis at 6 Ma and Australopithecus and Paranthropus in the later Pliocene is especially pronounced... Although among the early hominin taxa, Orrorin has the smallest Mahalanobis D2 distance from the modern human centroid, BAR 1002’00 is much closer to all early hominin taxa than to the modern human centroid in shape space. The external morphology of O. tugenensis provides no indications of differences in bipedal gait compared to Australopithecus or Paranthropus. This suggests that the pattern of bipedal gait characteristic of australopiths evolved very early in the human lineage, and perhaps they were also the characteristics of the first bipedal hominins. This form of bipedalism appears to have persisted as a successful locomotor strategy for as long as 4 million years. Additional lower limb fossils from the late Miocene and early Pliocene will be needed to test this hypothesis.The similarity between O. tugenensis and australopith femora weakens support for scenarios in which O. tugenesis is ancestral to Homo to the exclusion of A. afarensis. Instead, the overall primitive hominin morphology of the O. tugenensis femur, along with primitive dental anatomy, is consistent with the more parsimonious hypothesis that it is a basal member of the hominin clade. In sum, the comparative biomechanical anatomy of O. tugenensis femora suggests that O. tugenensis is a basal hominin adapted to bipedalism, and current evidence suggests that an Australopithecus-like bipedal morphology evolved early in the hominin clade and persisted successfully for most of human evolutionary history.[9] (Emphasis mine)
It would definitely be premature
to declare Orrorin tugenensis
directly on the line leading to Homo
sapiens, but it is eminently reasonable to assume that it is a primitive
hominin close to the time when the lines leading to Homo and Pan diverged,
and note that it is a splendid example of what special creationists claim does
not exist – a transitional fossil.
Sahelanthropus
tchadensis
Sahelanthropus
tchadensis is a late Miocene possible hominin located in the deserts of
Chad in 2002 and has been dated to around seven million years ago. The date
alone puts it on the early side of the possible time of the common ancestor as
determined by genetic studies. It is fair to say that there is no firm
consensus about whether it predates the Homo / Pan divergence, post-dates
the divergence and is on the Homo side of the divergence, or may even be
one of the earliest fossil non-human apes. Whatever it is, (remembering the
analogy of trying to work out what your grandfather looked like by looking at
photographs of his cousins, siblings and uncles) it is a valuable find.
The paper announcing its
discovery states that:
Sahelanthropus has several derived hominid features, including small, apically worn canines—which indicate a probable non-honing C–P3 complex—and intermediate postcanine enamel thickness. Several aspects of the basicranium (length, horizontal orientation, anterior position of the foramen magnum) and face (markedly reduced subnasal prognathism with no canine diastema, large continuous supraorbital torus) are similar to later hominids including Kenyanthropus and Homo. All these anatomical features indicate that Sahelanthropus belongs to the hominid clade.
In many other respects, however, Sahelanthropus exhibits a suite of primitive features including small brain size, a truncated triangular basioccipital bone, and the petrous portion of the temporal bone oriented 60° to the bicarotid chord. The observed mosaic of primitive and derived characters evident in Sahelanthropus indicates its phylogenetic position as a hominid close to the last common ancestor of humans and chimpanzees.[10]
The fossil remains are shown
below:

One problem palaeontologists face
is that the fossils are not infrequently deformed. This was the case with Sahelanthropus,
which makes interpretation difficult. Five years ago, the fossil was scanned,
and digitally reconstructed[11] in an
attempt to correct this deformation. The results appeared to support the theory
that it may have been very close to the common ancestor of humans and chimps:
The reconstruction in Fig. 2 is therefore a robust estimate of the cranial form of TM 266-01-60-1 that supports most of the details originally described. However, several features differ notably from those of the original specimen: the cranium as a whole is wider, the occipital contour is rounder sagittally, the nuchal plane is oriented more horizontally, the orbits are larger and more circular, and the face is superoinferiorly taller...The changes evident in the TM 266 reconstruction highlight its unique morphology and confirm several derived features shared with later hominids such as a relatively vertical face with an anteroposteriorly short premaxilla; an anteriorly-positioned foramen magnum linked to a relatively short basioccipital; a relatively flat, large, and horizontally-oriented nuchal plane; and downward lipping of the nuchal crest. These features, together with other dental features (see refs 1, 2), support the conclusion that Sahelanthropus is a hominid (contra Wolpoff et al).[12]
Of course, without post-cranial
data such as lower limbs, it is impossible to determine confidently that a
species known only by a cranium is bipedal or not. It is possible to infer it
however, using details in the skulls of bipeds such as the position of the
foramen magnum (the hole at the skull base through which the spinal cord exits)
as well as the orbital plane (the line from the top of the eye socket to the
bottom of the socket). In bipeds, the foramen magnum is located more towards
the bottom, while the orbital plane is perpendicular to the ground. The authors
of the paper detailing this reconstruction stated:
Finally, the TM 266 reconstruction permits an assessment of the hypothesis that Sahelanthropus was a biped, an important feature of Pliocene hominids and possibly several Late Miocene hominids. Unequivocal evidence for bipedalism is difficult to obtain from the cranium, but several lines of evidence suggest that TM 266-01-60-1 might have been bipedal. Despite substantial differences in neck orientation, humans and non-human primates tend to locomote with their orbital planes (the line joining the superior and inferior margins of the orbits) approximately perpendicular to the ground. In addition, primates orient the upper cervical vertebrae approximately perpendicular to the plane of the foramen magnum, and with only a limited range (about 10o) of flexion and extension possible at the cranio-cervical joint. The combined effect of these angular constraints is that the angle between the foramen magnum and the orbital plane (Fig. 4) is nearly perpendicular in Homo sapiens (103.2+/- 6.9o, n=23) but more acutely angled in Pan troglodytes (63.7+/-6.2o, n=20), and other species with more pronograde postures. The foramen magnum angle relative to the orbital plane in the TM 266 reconstruction is 95o, similar to that in humans and later bipedal hominids such as Australopithecus afarensis (AL 444-2) and A. africanus (Sts 5)13,17. TM 266-01-60-1 as a quadruped would require an unusually extended angle of the neck relative to the plane of the foramen magnum.[13] (Emphasis mine)
This reconstruction helped firm
support for the idea that it was a hominid. Building on this data, another
paper stated:
The hypothesis that TM 266-01-60-1 is an ape has been once again refuted by the recent virtual reconstruction , which confirms the presence of many hominid features in S. tchadensis and indicates that it was likely some kind of biped. However, questions remain about the species' systematic relationship to extant apes, other known Late Miocene hominids, and later hominids from the Pliocene. Although we do not believe it yet possible to resolve reliably the phylogenetic position of S. tchadensis and other early hominids, a first step is to consider in what ways the TM 266-01-60-1 cranium shares primitive features with extant apes and derived features with various known hominids. Thus, we assess here the morphological similarities and differences of the reconstructed TM 266-01-60-1 cranium with ape and other hominid taxa by using geometric morphometric methods along with comparative quantitative data on variables that reflect key similarities and differences in hominid craniofacial morphology.
The TM 266-01-60-1 reconstruction exhibits several widely recognized hominid synapomorphies, including a long, flat nuchal plane that is more horizontally oriented than in African apes (contra ref. 4); a shortened basioccipital; and a more anteriorly positioned foramen magnum... In addition, as discussed in Brunet et al., the dentition of TM 266-01-60-1 and referred specimens resembles those of Ardipithecus and later hominids compared with Pan in several respects such as reduced canines, a C/P3 nonhoning complex, and postcanine teeth with thicker enamel. TM 266-01-60-1 also shares several derived cranial features with later hominids, especially in the posterior cranial vault and the cranial base... However, as noted above, some aspects of the overall facial structure of TM 266-01-60-1 are unlike those of any known published Australopithecus, including AL 444-2 (A. afarensis). Several of these features, such as subnasal height, palatal protrusion, and relative upper facial projection...may be derived differently than those in Australopithecus. Others are probably either primitive or convergent with Homo. For example, the upper and middle portions of the face in TM 266-01-60-1 are similar in width, as in Pan and Homo, but unlike the hexagonal-shaped configuration in Australopithecus, in which the midface is wider than the upper face because of masticatory-related expansion of the zygomatic region.
Although Sahelanthropus tchadensis is clearly a hominid, its complex mosaic of features poses some interesting systematic questions about early hominid evolution that can be resolved only with more data of four types. First, more information is needed to compare Orrorin, Ardipithecus, and Sahelanthropus in detail. Second, additional hominid fossil material is needed from Late Miocene deposits in different parts of Africa. Third, we need a better understanding of the developmental and functional bases of cranial morphology that are necessary to hypothesize phylogenetically informative characters. Fourth, we particularly need a better understanding of the vertebrate and hominid biogeographical relationships through Africa during the upper Miocene.[14]
The authors placed Sahelanthropus tchadensis on a family
tree:

Note that it is placed before the
Australopithecus / Homo split, placing this fossil very close to
the human / chimp common ancestor. However, this position (as I mentioned
earlier) is opposed by some fairly respected anthropologists. In a 2006 paper,
Milford Wolpoff et al[15]
re-examined the data, and came up with a different conclusion.
This enigmatic Miocene species mixes characteristics of some australopithecines and some portions of the Homo clade such as “Homo erectus.” Yet, the hominid interpretation was not without objection (Wood, 2002) because TM 266 has also been described as a hominoid ape (Cela-Conde & Ayala, 2003; Wolpoff et al., 2002). The ape interpretation could mean that Sahelanthropus was uniquely ancestral to a living ape, or that the species was an extinct related lineage that diverged before the hominids, or that it is close to or actually the last common ancestor of hominids and chimpanzees.[16]
What is not in doubt is
that this species is transitional. However, as Wolpoff notes, it could be
ancestral to living apes, close to the Homo / Chimp common ancestor or even an
extinct line. After a fairly rigorous analysis of the data, they concluded:
Sahelanthropus was an ape living in an environment later abandoned by apes but subsequently inhabited by australopithecine species. Both TM 266 and the later australopithecines reflect the masticatory adaptations of earlier Miocene apes, and differ from later African apes in many of these adaptations. This creates the potential for significant homoplasies, and combined with the consequences of a limited fossil record, makes assessment of phylogeny problematic. Yet, it is a highly significant discovery; both because it evidences a unique pattern of homoplasy, and perhaps mostly because of the insight it might give for understanding the ancestral condition before the hominid-chimpanzee split.[17] (Emphasis mine)
However the authors of this paper
did not dismiss this fossil as having no bearing on evolution:
So every discovery counts. Sahelanthropus may be an early member of the gorilla clade, as two of us (B.S., M.P.) have suggested, or it may be closely related to the chimpanzee clade, or to the human-chimpanzee common ancestor (Wood, 2002), or perhaps most likely a member of an extinct closely related clade. In any of these cases, Sahelanthropus tends to confirm the notion (Wolpoff, 1982) that the ancestral condition for hominids, and for extant African apes, is much more like the Miocene ape condition than like living chimpanzees. Only if it turns out that Sahelanthropus represents a more distantly related hominoid lineage would it fail to be informative about the ancestral condition for these lineages, and address the issue of whether the chimpanzee is a reasonable model for this last common ancestor.[18] (Emphasis mine)
In short, the jury is out (though
for what it is worth I suspect Wolpoff may be correct, unless we can find
post-cranial data that definitely support bipedality). However, as we can see,
those who argue for and against it being ancestral to our line agree that it is
transitional, and of significance to the issue of human evolution.
Ardipithecus ramidus
The first Ardipithecus ramidus
fossils were found in 1992-1993 in the Middle Awash valley in Ethiopia, with
further fragments unearthed in 1994. It was formally named later year. Further
research in the early years of this century unearthed further evidence. Last
year, after some years of extensive research, 11 papers were published in
science detailing the results of that field work and subsequent analyis.
Now, before I go on any further,
I need to point out that Ardipithecus ramidus is not the common ancestor
since it is too young based on the currently accepted time for the divergence
of the lines leading to the Homo and Pan lineages based on
molecular studies. It however is still a significant find:
This remarkably rare skeleton is not the oldest putative hominin, but it is by far the most complete of the earliest specimens. It includes most of the skull and teeth, as well as the pelvis, hands, and feet—parts that the authors say reveal an "intermediate" form of upright walking, considered a hallmark of hominins. "We thought Lucy was the find of the century but, in retrospect, it isn't," says paleoanthropologist Andrew Hill of Yale University. "It's worth the wait."
To some researchers' surprise, the female skeleton doesn't look much like a chimpanzee, gorilla, or any of our closest living primate relatives. Even though this species probably lived soon after the dawn of humankind, it was not transitional between African apes and humans. "We have seen the ancestor, and it is not a chimpanzee," says paleoanthropologist Tim White of the University of California, Berkeley, co-director of the Middle Awash research group, which discovered and analyzed the fossils.
Instead, the skeleton and pieces of at least 35 additional individuals of Ar. ramidus reveal a new type of early hominin that was neither chimpanzee nor human. Although the team suspects that Ar. ramidus may have given rise to Lucy's genus, Australopithecus, the fossils "show for the first time that there is some new evolutionary grade of hominid that is not Australopithecus, that is not Homo," says paleontologist Michel Brunet of the College de France in Paris.[19]

Artist's
reconstructions show how Ardi's skeleton, muscles, and body looked and how she
would have moved on top of branches.
Credits: Illustrations © 2009, J. H. Matternes

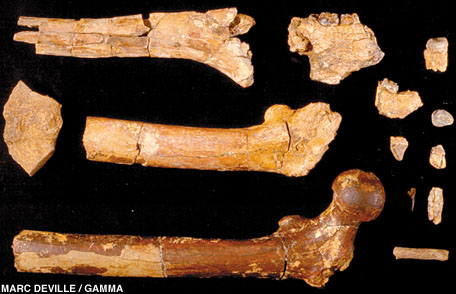
Fossil remains of Ardipithecus
ramidus.
Credit: Copyright T. White, 2008
Ardipithecus ramidus and
the likely post-cranial skeleton of the common ancestor
As I mentioned before, Ardipithecus
ramidus is too young to be the common ancestor, but it has a lot to say
about it. In so doing, it has invalidated hypotheses about this last common
ancestor:
The picture emerging from Ar. ramidus is that this last common ancestor had limb proportions more like those of monkeys than apes. Its feet functioned only partly like those of apes and much more like those of living monkeys and early apes such as Proconsul (which lived more than 15 million years ago). Its lower back was mobile and probably had six lumbar vertebrae rather than the three to four seen in the stiff backs of African apes. Its hand was unpredictably unique: Not only was its thumb musculature robust, unlike that of an ape, but its midcarpal joint (in the wrist) allowed the wrist to bend backward to a great degree, enhancing its ability to move along tree branches on its palms. None of the changes that apes have evolved to stiffen their hands for suspension and vertical climbing were present, so its locomotion did not resemble that of any living ape.
The hominid descendant of the last common ancestor we shared with chimpanzees (the CLCA), Ardipithecus, became a biped by modifying its upper pelvis without abandoning its grasping big toe. It was therefore an unpredicted and odd mosaic. It appears, unlike Au. afarensis, to have occupied the basal adaptive plateau of hominid natural history. It is so rife with anatomical surprises that no one could have imagined it without direct fossil evidence.[20]
Suffice it to say that Ardipithecus
is a transitional form: it is a biped with limb adaptations allowing it to
move in the trees. This is not simply another ape. So, where does Ardipithecus
possibly fit into the evolutionary tree? The diagram below shows where the
current data place it:

Cladogram
adding Ar. ramidus to images of
gorilla, chimpanzee, and human, taken from the frontispiece of Evidence as to
Man's Place in Nature, by Thomas H. Huxley (London, 1863) (with the positions
of Gorilla and Pan reversed to reflect current genetic data).
Numerous details of the Ar. ramidus skeleton confirm that extant African
apes do not much resemble our last common ancestor(s) with them. Credit:
Illustration of Ar. ramidus, copyright J. H. Matternes
Overall Summary.
Ardipithecus ramidus once again is not the common ancestor, but it gives us a very good idea of what the common ancestor may have been like. In so doing, it has negated some previous hypotheses about what that common ancestor would have been like - it would have looked more hominid-like than modern ape-like in many features. The authors note this:
"The most popular reconstructions of human evolution during the past century rested on the presumption that the behaviors of the earliest hominids were related to (or even natural amplifications of) behaviors observed in these living great apes...
"Ardipithecus ramidus nullifies these presumptions as it shows that the anatomy of living African apes is not primitive but instead has evolved specifically within extant ape lineages. The anatomy and behavior of early hominids are therefore unlikely to represent simple amplifications of those shared with modern apes. Instead, Ar. ramidus preserves some of the ancestral characteristics of the last common ancestor with much greater fidelity than do living African apes. Two obvious exceptions are its ability to walk upright and the absence of the large projecting canine tooth in males, derived features that Ardipithecus shares with all later hominids.
"Ar. ramidus illuminates our own origins because it clarifies our relationship to Australopithecus. For example, the enlarged rear teeth of Australopithecus have long been viewed as adaptations to a rough, abrasive diet. This has led to speculation that canine teeth might have become smaller simply to accommodate the emergence of these other enlarged teeth, or that the importance of canine teeth in displays of male-to-male aggression waned with the development of weapons.
"Ar. ramidus negates such hypotheses because it demonstrates that small canines occurred in hominids long before any of the dental modifications of Australopithecus or the use of stone tools. The loss of large canine teeth in males must have occurred within the context of a generalized, nonspecialized diet. Comparisons of the Ar. radius dentition with those of all other higher primates indicate that the species retained virtually no anatomical correlates of male-to-male conflict. Consistent with a diminished role of such agonism, the body size of Ar. ramidus males was only slightly larger than that of females.
"The discovery of Ar. ramidus also requires rejection of theories that presume a chimpanzee- or gorilla-like ancestor to explain habitual upright walking. Ar. ramidus was fully capable of bipedality and had evolved a substantially modified pelvis and foot with which to walk upright. At the same time, it preserved the ability to maneuver in trees, because it maintained a grasping big toe and a powerful hip and thigh musculature. Because upright walking provided no energy advantage for Ar. ramidus (it lacked many of the adaptations evolved in later hominids such as Australopithecus), reproductive success must have been central to its evolution in early hominids."[21]
This will not be the final word
on the subject, and details may change with further fossil evidence and/or
analysis of fossil data White has unearthed. However, this is an amazing find,
which does cast significant light on what the common ancestor would have been
like.
Appendix
The following covers Ardipithecus
ramidus in more detail for those interested.
Details of Hand Anatomy
"The thumb metacarpal of
ARA-VP-6/500 was more aptly proportioned for manual grasping than are those of
extant apes...
"Ar. ramidus greatly illuminates the natural history of the thumb in higher primates. Its robusticity in hominids, while certainly enhanced during the past 3 million years, is nevertheless at least partially primitive. In contrast, in taxa adapted to vertical climbing and suspension, lengthening of the palm has become so dominant as to eclipse some of the thumb’s function, a condition that has reached its apogee in Ateles and, to a lesser extent, large-bodied extant apes...
"Additional relevant hand anatomy leads to the same conclusions. Ar. ramidus is the only hominid fossil thus far recovered with a metacarpal head reminiscent of the metacarpophalangeal (MP) joint structure seen in many Miocene hominoids [such as Equatorius, Proconsul, Dryopithecus, and Pierolapithecus]. The collateral ligament facets in these taxa colocate with deep symmetric invaginations of the metacarpal head’s dorsum. This morphology is typical of Old World monkeys and is thereby associable with substantial dorsiflexion of the MP joint, an obvious manifestation of their palmigrady. The trait is only moderately expressed in Oreopithecus. Modern human and orangutan MP joints are substantially less constricted, and neither taxon exhibits appreciable locomotor-related MP dorsiflexion.[22]
Significance of hands for
common ancestor locomotion
"The primitive metacarpal
head morphology within the overall primitive hand anatomy of Ar. ramidus
carries obvious implications for reconstruction of GLCA/CLCA locomotion. The
unique combination of marked midcarpal mobility, ulnar withdrawal, and moderate
MP dorsiflexion in Ar. ramidus, probably mostly primitive retentions, implies
that the GLCA/CLCA locomotor pattern was also characterized by some form of
arboreal palmigrade quadrupedality, unlike that in any extant descendant great
ape.”[23]
Implications of the feet of Ardipithecus
ramidus for the common ancestor
"In summary, a comparison
of the pedal proportions of Ar. ramidus and the extant African apes
suggests that the GLCA/CLCA hindlimb remained dominant for body mass support
during bridging and arboreal clambering, to the extent that it later proved
permissive to bipedality in transitionally terrestrial hominids.”[24]
Details of the pelvis and
implications for the common ancestor
"Pelvic structure
indicates that Ar. ramidus retained a primitive spine. Its iliac and
acetabular regions establish not only that it was habitually bipedal when
terrestrial, but also that this was achieved by combining situational anterior
pelvic tilt to accentuate substantial lordosis during upright walking . Such
rotation placed the still partially primitive anterior gluteal musculature into
a position of functional abduction for single support stabilization. In
contrast to Ar. ramidus, Au. afarensis is known to have exhibited highly
evolved mechanisms of hip abduction, confirmed by the distinctly stereotypic
trabecular profile of its femoral neck.
"The Ar. ramidus pelvis retained other elements in common with extant African apes (and presumably the GLCA/CLCA). These include a long, expansive and rugose ischial region and shorter pubic rami (but not a long pubic corpus). The species’ highly flexible lower lumbar column, coupled with its narrower interacetabular distance, still must have provided a moderately reflexive hindlimb for arboreal climbing. Not until hominids became habitually terrestrial bipeds with broad interacetabular distances, reduced and angulated ischial tuberosities (possibly indicating hamstring deceleration of the hindlimb at heel strike), and extremely shortened, flared, and broadened ilia did they then exchange such flexibility for the much more rigid constraints of lower-limb stabilization that characterize Australopithecus.”[25]
"The Ar. ramidus pelvis retained other elements in common with extant African apes (and presumably the GLCA/CLCA). These include a long, expansive and rugose ischial region and shorter pubic rami (but not a long pubic corpus). The species’ highly flexible lower lumbar column, coupled with its narrower interacetabular distance, still must have provided a moderately reflexive hindlimb for arboreal climbing. Not until hominids became habitually terrestrial bipeds with broad interacetabular distances, reduced and angulated ischial tuberosities (possibly indicating hamstring deceleration of the hindlimb at heel strike), and extremely shortened, flared, and broadened ilia did they then exchange such flexibility for the much more rigid constraints of lower-limb stabilization that characterize Australopithecus.”[25]
Implications of postcranial
data for the common ancestor
"Therefore, we
hypothesize that Ar. ramidus retains much of the ancestral GLCA and CLCA
character states, i.e., those that relate to above branch quadrupedality. In
particular, contra Gorilla and Pan, the GLCA carpometacarpal,
midcarpal, radiocarpal, and ulnotrochlear joints must have lacked notable
adaptations to suspension and/or vertical climbing. The GLCA foot seems to have
been only partially modified for manual-like grasping. Its hindlimb remained
fully propulsive at its midtarsal and tarsometatarsal joints. Although its
shoulder joint must have been fully lateralized, its lumbar column nevertheless
was still long. Its limb proportions were still primitive (see earlier). If
body size was as large as in Ar. ramidus, it may have been too large for
habitual, unrestricted above-branch quadrupedality, but this remains uncertain.
Assuming considerable reliance on arboreal subsistence, it is likely that body
mass did not exceed 35 to 60 kg [i.e., combined probable range of Ar.
ramidus and 6 Ma Orrorin].
"The GLCA picture that emerges, therefore, is one of generalized, deliberate bridging with quadrupedal palmigrady and preference for large diameter substrates. This may have involved either suspension or vertical climbing, but without sufficient frequency to elicit morphological adaptations specific to these behaviors. It is likely that these hominoids ranged mostly in the lower canopy, and perhaps were even partially terrestrial. However, their mode of terrestrial locomotion remains unknown.
"The GLCA therefore represents a foundation for two adaptive paths. Gorilla and Pan independently specialized for both suspension and vertical climbing (and eventually knucklewalking). Gorillas might have acquired larger body size in relation to mixing higher-canopy frugivory with a more terrestrial herbaceous or folivorous dietary component. Lacking definitive fossil evidence, it is currently impossible to determine when the large body mass of Gorilla evolved, but it probably occurred in concert with its more herbaceous diet. The 10 Ma Chororapithecus, which shows incipient signs of Gorilla-like molar morphology, may be an early representative of the Gorilla clade. If so, then this clade’s shift toward increased body mass and terrestriality must have occurred early in its phyletic history.
"The other adaptive pathway retained palmar flexibility, with a short metacarpus that lacked notable syndesmotic restriction. This was combined with retention of an essentially rigid midtarsal joint that was insufficiently flexible to perform vertical climbing, but was fully satisfactory for less specialized careful climbing, clambering, and bridging. This is the hypothesized structure of the CLCA, from which Pan would have evolved a greater reliance on vertical climbing and suspension than occurred in the Gorilla clade, never reaching as large a body size.
"In contrast to Pan, the forebears of Ar. ramidus early in the hominid clade must have relied increasingly on lower arboreal resources and terrestrial zones, without being dependent on higher-canopy resources (such as ripe fruits). From the comparative evidence now available from Ar. ramidus and Pan dental anatomy and isotopes, we posit that the chimpanzee clade increasingly developed a preference for (or dependency on) ripe fruit frugivory, whereas hominids retained a more primitive dental complex adequate for the range of transitional arboreal/ terrestrial resources.[26]
The Foot of Ardipithecus
ramidus
The feet of modern apes and humans are different. While ours is rigid, and can act quite well as a lever during walking and running, the ape feet are less rigid and more effective for grasping - useful if you are mobilising through trees. It is not unreasonable to ask what the common ancestor's foot would have looked like - more like the chimpanzee foot, or more like ours. We now have an answer:
Hominid morphology has often been presumed to have evolved from ancestral morphotypes like those of extant African apes. Ar. ramidus now establishes that this was not the case. The hominid foot was instead derived from one substantially less specialized.[27]
In short, the foot of Ardipithecus
ramidus had a grasping great toe which would have helped it climb in trees,
while its four remaining toes were modified to make it suitable for upright
walking - it was a 'generalist' foot. A digitally rendered composite foot is
shown below:

Fig. 1 Digitally rendered composite foot of ARA-VP-6/500. (A) Plantar view. (B to D) Dorsal, medial, and anteromedial oblique views, respectively. Better-preserved elements from both sides were assembled as the left foot of Ar. ramidus. Mirror-imaged elements are the talus, cuboid, Mt2 shaft, and some phalanges. The intermediate and terminal phalanges are provisionally allocated to position and side. Note the anteroposteriorly strongly abducent first ray (Fig. 2), elongate cuboid (Fig. 3), and large os peroneal facet located more distolaterally than in Homo. Cuboids of African apes generally lack an os peroneum. Scale bars, 5 cm. Imagery is based on CT scans taken at 50- to 150-µm voxel resolution.
After a significant amount of technical detail, the authors interpret the significance of what they have found:
Ar. ramidus is the only known hominid with an abducent great toe. Its foot, along with other postcranial elements, indicates that the Late Miocene hominid precursors of Ar. ramidus practiced mixed arboreal and terrestrial locomotion during which the lateral forefoot became extensively adapted to upright walking, even as the medial forefoot retained adaptations for arboreal exploitation.[28]
In other words, it is quite
likely that the common ancestor (the Late Miocene hominid precursors) had a
foot whose lateral four toes were modified to permit upright walking, while the
medial forefoot allowed grasping. To date, it is the only hominid with a great
toe that allows grasping. This makes sense for a common ancestor, since this
means after the divergence of the Homo and Pan lines, the line
leading to us would become more and more suited to walking, and lose the need
for a grasping toe. Conversely, the line leading to chimpanzees and bonobos
would retain the grasping toe as they maintained a largely arboreal
environment.
We do see evidence of this
evolution towards more advanced bipedality in later hominids such as Australopithecus
afarensis:
Indeed, by the time of emergence of Au. afarensis, hominids had evolved substantially more advanced adaptations to bipedality than were present in Ardipithecus. In the former, the knee had become tibial dominant with accentuated valgus (exceeding even that of modern humans). Hip abduction had been established with a human-like distribution of proximal femoral cortical and trabecular bone. Moreover, in all known subsequent hominids, the more posterior location and elevation of the os peroneum facet on the cuboid [direct selection acting on morphogenetic fields; type 1] signals the presence of longitudinal and transverse arches, and thereby the addition of the transverse axis of fulcrumation (fig. S12). The facet’s position in the OH-8 cuboid is virtually human, as is the length of its calcaneal process. Doming and the simpler unnotched dorsal surface of the Mt1 head characterize both Au. afarensis (A.L. 333-21) and Au. africanus, confirming an immobile first ray with fundamentally human-like propulsion during toe-off.[29]
Having such a generalised foot
was critical for our evolution as bipeds. As the authors point out in their
summary, " If our last common ancestor with the chimpanzee had not
retained such an unspecialized foot, perhaps upright walking might never have
evolved in the first place."[30]
The Pelvis and Femur of Ardipithecus
Ramidus - Adaptations for upright walking
Our pelvis is of great significance in our bipedal gait - it helps up keep our balance on one lower limb while walking. If you look at chimps walking, the side of the pelvis to which the leg that is swinging forward is attached slumps down. That does not make for a very useful gait. Chimps and bonobos are our closest relatives, but they don't have a pelvic girdle like ours. The fossil record to date has not shed much light on the evolution of the early hominid pelvis. Ardipithecus ramidus may change this. From the author's summary:
Until now, the fossil record has told us little about when and how the early hominid pelvis evolved. Even 3 to 4 million years ago (when our brains were still only slightly larger than those of chimpanzees), it had already undergone radical transformation. One of the oldest hominid pelves, that of Australopithecus afarensis (A.L. 288-1; “Lucy”), shows that her species had already evolved virtually all of the fundamental adaptations to bipedality. Even the kinetics of her hip joint were similar to ours. Although the human pelvis was later further reshaped, this was largely the result of our much enlarged birth canal.Ardipithecus ramidus now unveils how our skeleton became progressively modified for bipedality. Although the foot anatomy of Ar. ramidus shows that it was still climbing trees, on the ground it walked upright. Its pelvis is a mosaic that, although far from being chimpanzee-like, is still much more primitive than that of Australopithecus...
The gluteal muscles had been repositioned so that Ar. ramidus could walk without shifting its center of mass from side to side. This is made clear not only by the shape of its ilium, but by the appearance of a special growth site unique to hominids among all primates (the anterior inferior iliac spine). However, its lower pelvis was still almost entirely ape-like, presumably because it still had massive hindlimb muscles for active climbing.
Changes made in the upper pelvis rendered Ar. ramidus an effective upright walker. It could also run, but probably with less speed and efficiency than humans. Running would also have exposed it to injury because it lacked advanced mechanisms such as those that would allow it to decelerate its limbs or modulate collision forces at its heel. Australopithecus, which had given up its grasping foot and abandoned active climbing, had evolved a lower pelvis that allowed it to run and walk for considerable distances.
Ar. ramidus thus illuminates two critical adaptive transitions in human evolution. In the first, from the human-chimp last common ancestor to Ardipithecus, modifications produced a mosaic pelvis that was useful for both climbing and upright walking. In the second, from Ardipithecus to Australopithecus, modifications produced a pelvis and lower limb that facilitated more effective upright walking and running but that were no longer useful for climbing. Because climbing to feed, nest, and escape predators is vital to all nonhuman primates, both of these transitions would likely have been a response to intense natural selection.[31]
The mosaic
nature of the pelvis is shown in the reconstruction below:

The Ar. ramidus pelvis has a mosaic of characters for both bipedality and climbing. Left to right: Human, Au. afarensis (“Lucy”), Ar. ramidus, Pan (chimpanzee). The ischial surface is angled near its midpoint to face upward in Lucy and the human (blue double arrows), showing that their hamstrings have undergone transformation for advanced bipedality, whereas they are primitive in the chimpanzee and Ar. ramidus (blue arrows). All three hominid ilia are vertically short and horizontally broad, forming a greater sciatic notch (white arrows) that is absent in Pan. A novel growth site [the anterior inferior iliac spine (yellow arrows)] is also lacking in Pan.
The pelvic
bones and their reconstructions are shown below:

Fig. 1 Original and reconstructed os coxa of ARA-VP-6/500. (Left) Anterolateral and anteromedial views of the original, with a close-up of the AIIS. (Middle) CT scans of same views, except the bottom view is a close-up of the pubic symphyseal face, preserved in its entirety but damaged at its inferior extremity. It is like later hominids in its dorsoventral height and ovoid outline. (Right) Anterolateral and anteromedial views of the reconstructed os coxa (11th permutation) and of the entire pelvis using mirror reconstruction and a conjectural sacrum. Various permutations were attempted with respect to sacral breadth; the solution shown provides presumed functional minimal inlet and outlet dimensions for a hominoid of this individual’s body size (Fig. 2 and figs. S1 to S3). (Middle) Arrows in the CT images indicate major areas of distortion corrected by reconstruction. The entire iliac blade was bent anteromedially (single yellow arrow) (additional and corrective data were provided by right iliac fragment). Two green arrows indicate primary foci of subduction of three largely intact segments of the superior pubic ramus; obvious overlapping allowed accurate restoration of original length (compare with reconstruction). The acetabulum was separated into two halves (total height of the exploded acetabulum is indicated by white arrows), with substantial intervening matrix infill. This separation greatly elongates its unrestored appearance. The two halves were recompressed on the basis of a calculated rim circumference obtained through the summation of individual segments (a range of probable values is presented in discussions of metric parameters). Two red arrows mark the inferior edge of the intact acetabular rim and the superior edge of the (missing) surface of the ischial tuberosity. These provide a minimum ischial length. Various additional dimensions were corrected by means of surface metrics. The breadth of the iliac fossa was intact from the AIIS to the lateral edge of the auricular surface; the degree of individual fragment separation was assessable from surface observation and CT scan data as indicated. Areas of particular anatomical importance include the protuberant sigmoid AIIS and sagittally oriented iliac isthmus, typical of later hominids, and the notably short, ovoid, pubic symphyseal face. Major metrics and angles are provided in Fig. 3. CT scans were taken at 300-µm voxel resolution on the University Museum, the University of Tokyo, micro-CT system [TX-225 Actis (Tesco, Tokyo)] and processed with the software Analyze 6.0 (Mayo Clinic, Rochester, MN) and Rapidform 2004/2006 (Inus Technology, Seoul).
Ilium, Ischium and pubis
Examination of these bones show clear evidence of transitional features:
"The protuberant and anteriorly positioned AIIS is associated with a broad, short, and sagittally disposed iliac isthmus. We define the isthmus as the constricted inferior portion of the iliac blade immediately superior to the acetabulum. These features are shared with later hominids, but in both modern and Early Miocene apes and monkeys the isthmus is markedly elongate, and the greater sciatic notch (GSN) angle is more obtuse. The Ar. ramidus GSN is intermediate between its counterpart in modern apes and those of later fossil hominids...
"In stark contrast to its distinctly hominid ilium, the preserved Ar. ramidus ischium is like that of African apes. Although the ischial tuberosity is not preserved, the ischium is intact from its typical concave surface flare just superior to the tuberosity to the inferior border of the acetabulum. Even this minimum preserved length of the superior ischial ramus is substantially longer than any known Australopithecus example. The pubis preserves an intact, superoinferiorly elongate body. However, the outline of the pubic symphyseal face is similar to that of Homo and A.L. 288.-1 and unlike its extreme dorsoventral elongation in African apes.[32]
"In stark contrast to its distinctly hominid ilium, the preserved Ar. ramidus ischium is like that of African apes. Although the ischial tuberosity is not preserved, the ischium is intact from its typical concave surface flare just superior to the tuberosity to the inferior border of the acetabulum (Fig. 1). Even this minimum preserved length of the superior ischial ramus is substantially longer than any known Australopithecus example. The pubis preserves an intact, superoinferiorly elongate body. However, the outline of the pubic symphyseal face is similar to that of Homo and A.L. 288.-1 and unlike its extreme dorsoventral elongation in African apes (Fig. 3A). This more ovoid shape may be a collateral pleiotropic manifestation...of the shortened iliac isthmus and elongated superior pubic ramus, especially in A.L. 288-1 (which exhibits both an unusually broad pubic face and greatly elongated pubic rami)."[33]
Pelvis form and function
Examination of the pelvis of Ardipithecus ramidus shows evidence of change
"The form and size of the AIIS in ARA-VP- 6/500, as well as its projection anterior to the acetabular margin, indicate that this structure had already begun to appear and mature via a novel physis. Isolation of the AIIS as a separate growth center, which is unique to hominids, was probably a consequence of its increased separation from the original iliac portion of the acetabular chondroepiphysis. This is because much of iliac broadening must occur at the triradial epiphysis (as well as the posterior iliac crest). An analogous phenomenon occurs in mammals with elongate femoral necks, in which accelerated growth in the region separating the presumptive greater trochanter and femoral head isolates these structures before separate ossification of each epiphysis.
"The emergence of a novel AIIS center of ossification, as seen in ARA-VP-6/500, attests to global modification of the entire pelvis. This is also demonstrated by the abbreviated craniocaudal length of the pubic symphyseal face. The anteroventral pubic surface is surmounted by a rugose pectineal line that is continuous with the nonperiosteal attachments of abductor brevis and gracilis muscles. This is a derived character in human females... It is only rarely present in great apes. There is no lateral displacement of this feature in Ar. ramidus as there is in human females (and partially in A.L. 288-1)...
"Given retention by Ar. ramidus of multiple primitive skeletal characters...its exceptionally derived ilium is striking. It implies an early adaptation to habitual terrestrial bipedality before any increase in the lumbar entrapment seen in the African apes, but after the lateral iliac expansion shared with them.”[34]
Conclusion
We see in the feet of Ardipithecus adaptations both for tree climbing and walking. What we see in the pelvis are clear signs of modification of a pelvis for walking:
The pelvis, femur, and preserved thoracic elements of Ar. ramidus establish that adaptations to upright walking in these regions were well established by 4.4 Ma, despite retention of a capacity for substantial arboreal locomotion. Ar. ramidus thus now provides evidence on the long-sought locomotor transition from arboreal life to habitual terrestrial bipedality. This evidence suggests that the transition took place in the absence of any of the characters that today substantially restrict upright walking in extant apes (particularly lumbar column abbreviation, trans-iliac space narrowing, and approximation of iliac crest and thorax, and the muscles that traverse this gap). As a consequence, explications of the emergence of bipedality based on observations made of African ape locomotion no longer constitute a useful paradigm.[35]
The forelimbs of Ardipithecus ramidus and their comparison to human
forelimbs
One defining feature of primates are highly mobile upper limbs with hands that are capable of grasping and manipulating objects. While chimps and gorillas can walk upright, they also knuckle walk - a trait we don't have since we no longer use our arms for propulsion. This raises the question: did the common ancestor knuckle walk (which meant that this trait was lost by the line leading to us after divergence from the common ancestor) or was its anatomy more generalised?
We now have an answer. In their summary, the authors state that:
"Ardipithecus hands were very different from those of African apes. Its wrist joints were not as stiff as those of apes, and the joints between their palms and fingers were much more flexible. Moreover, a large joint in the middle of the wrist (the midcarpal joint) was especially flexible, being even more mobile than our own. This would have allowed Ardipithecus to support nearly all of its body weight on its palms when moving along tree branches, so that it could move well forward of a supporting forelimb without first releasing its grip on a branch.
"This discovery ends years of speculation about the course of human evolution. Our ancestors’ hands differed profoundly from those of living great apes, and therefore the two must have substantially differed in the ways they climbed, fed, and nested. It is African apes who have evolved so extensively since we shared our last common ancestor, not humans or our immediate hominid ancestors. Hands of the earliest hominids were less ape-like than ours and quite different from those of any living form."[36]
Here's a
picture of the hand model:

Two views of the left hand of Ar. ramidus showing primitive features absent in specialized apes. (A) Short metacarpals; (B) lack of knuckle-walking grooves; (C) extended joint surface on fifth digit; (D) thumb more robust than in apes; (E) insertion gable for long flexor tendon (sometimes absent in apes); (F) hamate allows palm to flex; (G) simple wrist joints; (H) capitate head promotes strong palm flexion. Inset: lateral view of capitates of Pan, Ar. ramidus, and human (left to right). Dashed lines reflect a more palmar capitate head location for Ar. ramidus and humans, which allows a more flexible wrist in hominids.
Analysis of the hand bones does
not show the traits you would expect to see in knuckle walking:
"The medial metacarpals (Mc2 to Mc5) of Ar. ramidus are short compared with those of African apes. Their heads lack any traits associated with knuckle-walking, such as prominent ridges and/or grooves. Ar. ramidus metacarpal heads exhibit marked, proximally located, dorsal invaginations that are associated with their metacarpophalangeal (MP) joint’s collateral ligaments (reflecting routine hyperdorsiflexion at this joint), as do those of some Old and New World monkeys and some Miocene hominoids, including Proconsul...Such constrictions have become minimal in all extant hominoids but for different functional reasons... Ar. ramidus also lacks the expansion of the metacarpal heads that is typical of knuckle-walking apes...
"The first ray’s terminal phalanx exhibits a symmetrically constructed, rugose insertion gable for the flexor pollicis longus, in contrast to African apes in which this muscle’s tendon is reduced or absent."[37]
The hands of apes are actually
specialised for things like knuckle walking and tree climbing. Ardipithecus
had a more generalised hand which we have generally retained (though with
somewhat shorter fingers!)
"In summary, the hand of Ar. ramidus appears almost entirely primitive relative to the anatomical specializations seen in extant apes (for example, metacarpal elongation, elaboration of CJC articulations and ligaments, novel capitate geometry, reorientation of the scaphoid’s radial surface, enlargement of the radioscaphoid ligament, relative diminution of the first ray, etc.). Ar. ramidus establishes that these changes in the ape hand are independent specializations for arboreal access and terrestrial travel (vertical climbing, forelimb suspension, knuckle-walking) and were apparently never established in hominids, which retained a more generalized, substrate-conforming, grasping hand.”[38]
Finally, Ardipithecus
allows us to postulate that our hominid ancestors likely never were knuckle
walkers.
"The forelimb has played a definitive role in most chronicles of human evolution since Huxley’s and Keith’s original accounts. Most recent narratives of its anatomical and behavioral evolution have emphasized a heritage of suspensory locomotion, vertical climbing, and knuckle-walking in the common ancestors that humans shared with extant African apes.
"Encouraged by human and chimpanzee genetic similarity and cladistic analyses, such views have come to dominate recent explications of early hominid evolution, although alternative interpretations based on classical comparative anatomy have long differed. Ar. ramidus now permits resolution of these controversies. It indicates that, although cranially, dentally, and postcranially substantially more primitive than Australopithecus, these known Late Miocene to Early Pliocene hominids probably all lacked the numerous, apparently derived, forelimb features of extant African apes. The most probable hypothesis to explain these observations is that hominids never passed through adaptive stages that relied on suspension, vertical climbing, or knuckle-walking. Further fossil remains from the Late Miocene, including those before and after the African ape–hominid phyletic divergences, will test this hypothesis derived from our analysis of Ar. radius."[39]
Ardipithecus ramidus
Skull
It is somewhat uncommon to have complete hominid skulls, which is why complete skulls are quite prized. Obviously, the more complete skulls you have, the better you can track the evolution of brain, face and skull. Furthermore, one can infer details of diet and behaviour from skulls.
A reassonably complete - though crushed - skull was found, which meant that the bones needed by be scanned and digitally reassembled to reconstruct the skull. The key findings, to quote from the author summary:[40]
- It had a
small brain (300-350 cc), smaller than the Australopithecines (400 -550
cc), and around the size of bonobos and female chimpanzees
- Its face
was small, and did not have the large cheek bones that we see in
Australopithecines that were capable of grinding tough food
- It had a
projecting muzzle, similar to Sahelanthropus, giving it an ape-like
feel
- However,
its skull does not resemble that of chimps closely.
There are interesting comparisons
between its skull and that of Sahelanthropus:
Like Ar. ramidus, S. tchadensis had a brain that was less than 400 cm3 in size. It also resembled Ar. ramidus in having small nonsharpened canines. Details of the bottom of the skull show that both Ar. ramidus and Sahelanthropus had a short cranial base, a feature also shared with Australopithecus. Furthermore, we infer that the rear of the Ar. ramidus skull was downturned like that suggested for Sahelanthropus. These similarities confirm that Sahelanthropus was indeed a hominid, not an extinct ape.[41]
One of course recalls Wolpoff et
al and their caution about the status of Sahelanthropus. Ardipithecus
may well tip the basis in favour of Sahelanthropus being a hominid -
time will tell. Their conclusion:
These and an additional feature of the skull hint that, despite its small size, the brain of Ar. ramidus may have already begun to develop some aspects of later hominid-like form and function. The steep orientation of the bone on which the brain stem rests suggests that the base of the Ar. ramidus brain might have been more flexed than in apes. In Australopithecus, a flexed cranial base occurs together with expansion of the posterior parietal cortex, a part of the modern human brain involved in aspects of visual and spatial perception.[42]
The fragmented skull bones are
shown below:

Fig. 1 The fragmented skull of ARA-VP-6/500. (Upper panel) Identifiable pieces of the skull after limited refitting for digital and physical molding. (Lower panel) (A) ARA-VP-6/500-032, (B) micro-CT rendered image of the same, with cross-sectional locations of (C) and (D) indicated. Arrowheads in (A) denote the positions of (C) and (D).


(Right) Oblique and side views of a female chimpanzee (right) and the Ar. ramidus female reconstruction (left; the oblique view includes a separate mandible). (Left) Comparison of brain and tooth sizes (arrows) of chimps (Pan, blue), Ar. ramidus (red), and Australopithecus (green). Means are plotted except for individual Ar. ramidus and Au. afarensis cranial capacities. Canine unworn heights (bottom) are based on small samples, Ar. ramidus (females, n = 1; males, n = 3), Au. afarensis (n = 2), Pan (females, n = 19; males n = 11
[1]
Meade L.S. “Transforming Our Thinking About
Transitional Forms”
Evo Edu Outreach (2009) 2:310-314
[4]
See also the lay-accessible
series at BioLogos by palaeoanthropologist and Christian James Kidder.
[5]
Senut B et al "First
hominid from the Miocene (Lukeino Formation, Kenya)" Earth and
Planetary Sciences (2001) 332:137–144
[6]
ibid, p 139
[7]
ibid, p 142
[8]
. K. Galik et al "External
and Internal Morphology of the BAR 1002'00 Orrorin tugenensis Femur"
Science (2008) 305:1450-1453
[9]
Richmond BG, Jungers WL "Orrorin
tugenensis Femoral Morphology and the Evolution of Hominin Bipedalism"
Science (2008) 319:1662-64
[10]
Brunet M et al "A
new hominid from the Upper Miocene of Chad, Central Africa" Nature
(2002) 418:145-151
[11]
Zollikofer CPE et al "Virtual cranial
reconstruction of Sahelanthropus
tchadensis" Nature (2005) 434:755-759
[12]
ibid p 756
[13]
ibid, p 757-758
[14]
Guy F et al "Morphological
affinities of the Sahelanthropus
tchadensis (Late Miocene hominid from Chad) cranium" Proc Natl
Acad Sci USA (2005) 102:18836-18841
[15]
Wolpoff MH et al "An Ape
or the Ape: Is the Toumaï Cranium TM 266 a Hominid?" PaleoAnthropology
2006: 36−50
[16]
ibid, p 37
[17]
ibid, p47
[18]
ibid, p 48
[20]
C. O. Lovejoy et al. "The Great Divides: Ardipithecus
ramidus Reveals the Postcrania of Our Last Common Ancestors with African
Apes" Science (2009) 326:73. Author Summary)[21]
Lovejoy CO "Reexamining
Human Origins in Light of Ardipithecus ramidus" Science (2009) 326:74
[22]
C. O. Lovejoy et al. "The
Great Divides: Ardipithecus ramidus
Reveals the Postcrania of Our Last Common Ancestors with African Apes"
Science (2009) 326:73. Author Summary)
[25]
Lovejoy, p 103
[26]
loc cit
[27]
Lovejoy CO et al "Combining Prehension and
Propulsion: The foot of Ardipithecus ramidus" Science
(2009) 326:72
[28]
ibid, p 72e5
[29]
ibid, p 72e6
[30] ibid,
p 72
[31] .O. Lovejoy et al "The Pelvis and Femur of Ardipithecus ramidus: The Emergence of Upright Walking" Science (2009) 326:71, 71e1-71e6
[32] ibid, p 71e1
[33] loc cit
[34] ibid, p 71e3
[35] ibid, p 71e5
[36] Lovejoy CO et al "Careful Climbing in the Miocene: The Forelimbs of Ardipithecus ramidus and Humans Are Primitive" Science (2009) 326:70
[37] ibid, p 70e1
[38] ibid, p 70e5
[39] ibid, p 70e6-7
[40] Suwa G et al. "The Ardipithecus ramidus Skull and Its Implications for Hominid Origins" Science (2009) 326:68
[41] loc cit
[42] loc cit
[31] .O. Lovejoy et al "The Pelvis and Femur of Ardipithecus ramidus: The Emergence of Upright Walking" Science (2009) 326:71, 71e1-71e6
[32] ibid, p 71e1
[33] loc cit
[34] ibid, p 71e3
[35] ibid, p 71e5
[36] Lovejoy CO et al "Careful Climbing in the Miocene: The Forelimbs of Ardipithecus ramidus and Humans Are Primitive" Science (2009) 326:70
[37] ibid, p 70e1
[38] ibid, p 70e5
[39] ibid, p 70e6-7
[40] Suwa G et al. "The Ardipithecus ramidus Skull and Its Implications for Hominid Origins" Science (2009) 326:68
[41] loc cit
[42] loc cit


{kind=link}