Scientific errors in Christadelphian attacks on evolution: The Testimony magazine - 8
While our community indulges in science denialism by encouraging pseudoscientific nonsense such as YEC, and actively attacking evolution, we will continue to drive away scientifically literate young people. Comments such as this are hardly uncommon:
“My own eldest son has decided he cannot be baptized because he has seen the evidence for evolution with his own eyes, and our ecclesia will not tolerate discussion on the subject. Unlike some young people, he is too honest to say he doesn’t believe it, just so that he can ‘pass the test’ and be baptized.”
“I will be spending most of this semester studying common descent and evolution in first year biology, and have done so through DNA and cells so far. It really is fascinating and very undeniable. There's also a young Christo girl from [X ecclesia] in the subject, and I am interested to know what she's thinking.”
Unfortunately, The Testimony is actively encouraging science denialism while it continues to publish poorly reasoned material, such as John Watts' article [1] attacking common descent. Material such as this destroys our credibility in the eyes of young people and potential converts who recognise the overwhelming evidence for evolution.
The précis for Watts’ article immediately highlights the major flaws in his argument:
It is often argued that similarities between species support the theory of evolution. This article shows that this argument is not sound, since we should expect such similarities in a world created as described in the Bible.
Watts fails to differentiate between common descent and the modern synthetic theory – a common failing of creationists who routinely conflate the fact of common descent and the mechanisms postulated to explain it. Assuming he means the former, then Watts needs to show that common descent is not a reasonable explanation for the data. Furthermore, he needs to show that special creation is a better explanation. Given what we’ve seen of the data, it is difficult to see how special creation would account for shared ERV elements.
Watts claims:
A recent article claims to demonstrate beyond any doubt that apes and man had a common ancestor. The evidence used is drawn from cytology and molecular biology, considering similarities first in the physical structure of parts of the chromosomes of man and primates, and then in some of the base sequences of their DNA. It is another example of reasoning from comparative anatomy.
While it was not reasonable to expect Watts to go into specific technical details given the target audience of The Testimony and space constraints, he was obliged to accurately summarise the argument he is criticising. He did not done that. The argument is more than just similarity in chromosomal banding and base sequences. Rather, as we’ve seen, it stretches past homology and synteny to common genetic errors in related species which cannot credibly be explained either by chance insertion of retrotransposed element or by common design. Watts has failed to accurately summarise Finlay’s argument. At this point, the remainder of his article can be dismissed since it is merely an attack on a straw man version of Finlay’s paper.
In his opening paragraph, Watts dismissed the argument from comparative genomics as one that “relies more on ignorance than on knowledge.” The irony of course is that Watts has demonstrated that he is largely ignorant of contemporary developments in molecular biology and genomics. Apart from failing to accurately summarise Finlay’s argument (which is hardly Finlay’s idea, but represents the overwhelming consensus of professional molecular biologists working in the field) Watts does not even cite him once. Watts’ following paragraph does little to disabuse the informed reader of this belief:
Scientists, if they are honest, know that they operate in a fog of ignorance. Advances in un-derstanding are very slow and uncertain, and the ‘theories’ of today are usually little more than a consensus of opinions and never fit all the known facts.
The opening sentence is both patronising and gratuitous, as scientists freely acknowledge this fact. Nothing in science is ever regarded as final, but held provisionally pending further evidence. Watts is less than subtly trading on the lay perception of the ‘arrogant scientist’ to make his assertion that molecular biologists are prematurely declaring that the evidence supports common descent. Not only does that ignore the sheer weight and unambiguous nature of the evidence as shown earlier, it mischaracterises the scientific process. The evolutionary biologist TR Gregory reminds his readers that:
It is this insistence on repeated confirmation by data—either through direct observation or reliable inference—that makes a claim to “fact” so much stronger in science. However, as the NAS points out, “truth in science is never final, and what is accepted as a fact today may be modified or even discarded tomorrow”. Small-scale details are regularly revised as more precise observations are made, whereas well established facts of fundamental significance are very rarely overthrown, but in principle, no scientific fact of any magnitude is beyond revision or refutation. As a result, scientists must maintain a balance between the confidence that comes from reinforcing conclusions about the world with repeatable data and the understanding that absolute certainty is not something that the methods of science are able or intended to deliver. [2]
In particular, Watts demonstrates a poor understanding of what a scientific theory is. To call it a ‘consensus of opinions’ that ‘never fit all the known facts’ is a distortion at best. Gregory’s comment on science means by ‘theory’ is worth quoting if only to show the degree of Watts’ error:
The common and scientific definitions of “theory,” unlike of “fact,” are drastically different. In daily conversation, “theory” often implicitly indicates a lack of supporting data. Indeed, introducing a statement with “My theory is...” is usually akin to saying “I guess that...”, “I would speculate that...”, or “I believe but have not attempted to demonstrate that...”. By contrast, a theory in science, again following the definition given by the NAS, is “a well-substantiated explanation of some aspect of the natural world that can incorporate facts, laws, inferences, and tested hypotheses.” Science not only generates facts but seeks to explain them, and the interlocking and well-supported explanations for those facts are known as theories. Theories allow aspects of the natural world not only to be described, but to be understood. Far from being unsubstantiated speculations, theories are the ultimate goal of science. [3]
Watts continues his argument by claiming that:
The source of power of arguments like this one, based on chromosome architecture and base sequences, is derived from ignorance; they look similar, therefore they are related, and no one knows enough to be able to offer constructive criticism. Arguments from similarities invariably lose much of their plausibility when the things being compared are better understood.
The irony of Watts’ claim that such arguments are ‘derived from ignorance’ is palpable given that he has shown his ignorance of the subject he criticised, in claiming that it is based on “chromosome architecture and base sequences.” His argument is analogous to claiming that the evidence for plagiarism is simply based on paragraph structure and letter sequences, ignoring the fact that shared spelling errors in the same places in identical paragraphs is better explained by copying than by independent generation of the same mistake by the students.
Enough has been said earlier, so there is little point in elaborating this evidence further other than to point out that Watts needs to explain why an intelligent designer would choose a tiny subset of the coding space for cytochrome c genes and structure it in such a way as to simulate descent with modification, or create humans and chimpanzees with multiple shared ERV elements.
Watts continues:
How can anyone usefully comment on an argument based on visible patterns in chromosomes, objects whose structures and purposes are poorly understood? What can we usefully say about base sequences in DNA when over ninety-nine per cent of DNA is dismissed as ‘junk’, in other words as of unknown significance? Apparent similarities between species will always be seized on to support evolution.
Watts’ polemic betrays a poor understanding of the subject he criticises. If he had truly understood Finlay’s paper, he would have recognised that the most powerful evidence for common descent comes from shared retrotransposed elements, as these are unarguable evidence of insertion into the genome from another source, and the presence of shared identical retrotransposed elements is prima facie evidence of a retrotransposition event in a common ancestor of those species. Instead, he continues as if the strongest evidence is similarity of chromosomal banding. It isn’t. We’ve progressed well past karyotyping to genome sequencing and bioinformatics.
This less than comprehensive understanding of genomics is revealed when he asserts that chromosomes are “objects whose structures and purposes are poorly understood.” Modern science has sequenced many genomes, to the point where the field of comparative genomics has emerged. We can compare genes between species, construct phylogenetic trees based on the genetic data, compare syntenic blocks between species and by using this data reconstruct an ancestral genome for the species compared. In terms of genome structure, our knowledge is hardly poorly understood.
With respect to function, there is still much about which we are ignorant, but this is hardly a secret. Rather, understanding the function of the human genome is one of the hottest areas of research in molecular biology today. However, this does not alter the fact that we have a very good idea of the structure of the genomes of many organisms down to the base sequence level, and this as mentioned before unarguably shows evidence for common descent.
Let’s return for a moment to the example of how shared spelling errors in the same places in term papers are evidence of plagiarism, rather than independently occurring mistakes. If these term papers were in an abstruse subject such as the philosophy of biology, and three students had copied the papers of a fourth student who had copied the argument made in a paper found on-line in a term paper archive. Furthermore, this on-line term paper contained numerous spelling mistakes and grammatical errors. A layperson who read these papers would not understand much of the paper given her lack of training in the philosophy of science, but she would still be able to see evidence of shared spelling and grammar errors in identical places in the papers. Furthermore, if she performed an internet search and discovered the offending paragraph in the paper the fourth student had plagiarised, she would have enough evidence to conclude that wholesale plagiarism and copying had occurred, even if she did not know anything about the philosophy of science. The same applies here – what we do not know about comparative genomics does not make the overwhelming evidence for common descent from gene homology, synteny, shared pseudogenes and shared retrotransposed elements go away.
Watts further erodes the credibility of his argument when he asserts:
Even the use of the fossil ‘record’ is only an extension of the argument, since no transitional stages between species have been observed.
He demonstrates his ignorance of palaeontology by implying that the paucity of transitional fossils at species level is a problem for evolutionary biology. As the palaeontologist Stephen Jay Gould pointed out nearly 30 years ago:
Since we proposed punctuated equilibria to explain trends, it is infuriating to be quoted again and again by creationists -- whether through design or stupidity, I do not know -- as admitting that the fossil record includes no transitional forms. Transitional forms are generally lacking at the species level, but they are abundant between larger groups. [4]
Given that the overwhelming majority of dead organisms are consumed after death, fossilisation is an unlikely event. It is hardly special pleading to point out that the fossil record is fragmentary – we are fortunate to have enough fossils to document large scale evolutionary change at higher taxonomic levels. [5-8] However, there are examples of species-level transitional fossils in which the fossil record is so rich and detailed that we can see evolution at the species level. The vertebrate palaeontologist Donald Prothero points out that:
But there are circumstances where the fossil record is so complete that it is possible to say that "the fossils in this population represent the ancestors of this later population." [Prothero and Lazarus 1980] provided just such an example from the extraordinary fossil record of planktonic microfossils. In these unusual circumstances, we have deep-sea cores covering all of geologic time since the Jurassic for most of the world's oceans and every centimeter of sediment in most of these cores is filled with thousands of microfossils. With an extraordinarily dense and continuous record such as this, we really can say that we have sampled all the fossil populations that lived in the world's oceans and can establish which samples are most likely the ancestors of later populations." [9]
Prothero continues by giving specific examples of species-level transitional fossils on which he had worked earlier in his career:
From the ancestral form Pterocanium charybdeum allium...that lived 7 million years ago, we documented a complex pattern of divergence among different shapes. Some developed larger pores, more robust spines that flared out from the base, and lost the distinct "knob" of the cephalis at the top (Pterocanium audax). Another lineage developed a more cylindrical, boxy shape with distinct "shoulders." (Pterocanium prismatium, an important index fossil for the Pliocene). Another lineage developed huge flaring spines and shrank the size of the thorax to a little ball (Pterocanium korotnevi). Two lineages that had been distinct for millions of years (Pterocanium praetextum and Pterocanium charybdeum trilobum) appeared to hybridize during the late Pleistocene...[It] was possible to measure many different features, such as the length of the thorax, and see the gradual shift of sizes in each population spanning 7 million years.
This is just one example of evolutionary changes among the radiolaria, and there are hundreds more that could be cited...Let us look at just one more classic example, probably the most extreme change in morphology ever documented in the fossil record. If you look at the samples of microfossils from the middle Eocene (50 million years ago), you will find distinctive spongy ball-shaped radiolarians known as Lithocyclia ocellus...As you trace the spongy balls up through the sediments spanning millions of years, you see them gradually lose their spongy outer layers and develop into a small nucleus with four spongy arms (Lithocyclia aristotelis), then three arms (Lithocyclia angusta), and finally reduced to two arms forming a spindle-like shape (Cannartus tubarius.) The Cannartus lineage then gradually develops a "waist" on the central sphere, then the arms get shorter and thicker, and finally, they split into two lineages...If you look at the two extremes (a spongy sphere turning into a spindle-shaped shell with multiple caps), you could never imagine that they are closely related - yet I have looked at the slides from those cores and seen the gradual transition from one extreme to the other with my own eyes. (Emphasis mine) [10]
Not only do we have numerous transitional fossils documenting large-scale evolutionary change at higher taxonomic levels, we also have in the microfossil record evidence of species-level evolutionary change. In claiming that there are no transitional fossils between species, Watts is wrong. More to the point, he has demonstrated a fundamental ignorance of palaeontology, and that destroys his credibility in this area.
Despite the fact that Watts is ostensibly rebutting the claim that comparative genomics provides overwhelming evidence for common descent, not only has he not made any substantive comment on the single article he cites, he spends much of his article claiming that similarity in morphology is a function of design constraints and efficiency:
In any efficient design, the end use determines the form of the structure. Most steam, petrol and diesel engines use some sort of crank to convert the linear reciprocating movement of a piston in a cylinder to a circular motion that can turn a wheel. They share this structure because it has proved an efficient way to achieve a particular end.
Watts then argues “It may be that the ‘similarities’ in comparative anatomy reflect a similar constraint.” However, the next example he gives not only fails to support his claim, but in fact is strong evidence for common descent.
The eye in the octopus is very similar to that of the human, but no evolutionist invokes the similarity to prove evolutionary relationships
One would hardly expect an evolutionary biologist to say that, since humans and the various species of octopus are regarded by biologists as being very distantly related. In fact, they’re in completely different phylums! Humans are chordates, while octopuses are molluscs, and evolutionary biologists state that their last common ancestor lived over 700 million years ago.[11]
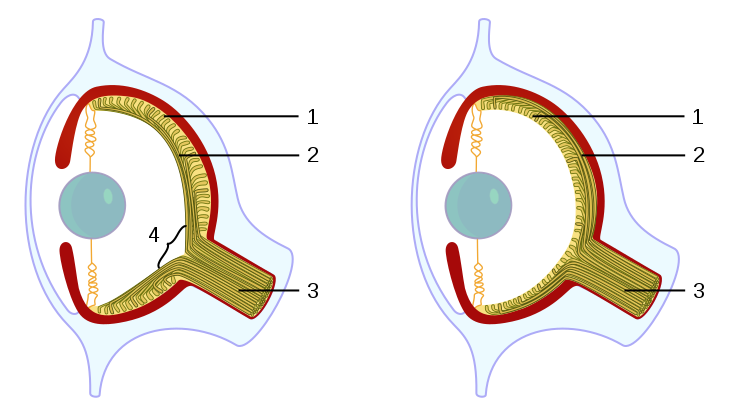 |
| Vertebrates and octopuses developed the camera eye independently. In the vertebrate version the nerve fibers pass in front of the retina, and there is a blind spot where the nerves pass through the retina. In the vertebrate example, 4 represents the blind spot, which is notably absent from the octopus eye. In vertebrates, 1 represents the retina and 2 is the nerve fibers, including the optic nerve (3), whereas in the octopus eye, 1 and 2 represent the nerve fibers and retina respectively. (Source: Wikipedia) |
Furthermore, Watts makes another mistake by claiming that octopus and human eyes are very similar. In one fundamental area, they are radically different. In octopuses, the light sensing cells in the retina point towards the light, whereas in humans, they point backwards away from the light. This results in a number of problems [12] with the human eye:
- As the nerve fibres from the light sensing cells need to leave the eye, there is a hole in the retina through which they pass, known as the blind spot. If the light sensing cells pointed towards the light, the nerve fibres would be behind them, and would be able to exit the eye without the need for a hole in the retina. Cephalopods such as the octopus in fact do not have a blind spot as their light cells point towards light.
- As the light sensing cells point away from the light in humans, this means the blood vessels are between them and the light. The condition diabetic retinopathy causes these blood vessels to grow and multiply abnormally, obscuring vision to the point where laser treatment to burn these excess blood vessels away is often needed. This condition is a direct result of the design of the human eye.
- The back-to-front design of the human eye makes it susceptible to retinal detachment. If the eye had a front to back design, the nerve fibres would anchor the retina firmly to the eye and retinal detachment would not occur to anywhere near the frequency it does in humans.
- In order to compensate for the back-to-front design of the human eye, in one part of the eye called the macula, the nerves and blood vessels are pushed away to allow an increase in photocell density and increase visual acuity. Not only would this not be needed if the human eye followed the octopus design with the blood vessels and nerve fibres behind the light sensing cells, it also leaves humans susceptible to macular degeneration. If the macula degenerates, then human visual acuity can plummet dramatically. At worst, it can leave people legally blind.
The arrangement of the extraocular muscles—the muscles that move the eyes—is also difficult to explain without appealing to evolutionary contingency. There are more muscles than are minimally necessary and yet there is no functional redundancy. In order to move a sphere in any direction, only three muscles would be necessary, evenly spaced like the legs of a tripod. The human eye has six—the superior, inferior, lateral, and medial rectus, and the superior and inferior oblique. And yet, despite the extra three muscles, the loss of function of any one muscle causes an impairment of eye movement and results in double vision or displaced vision. A more frugal design with only three muscles would be more efficient and less prone to malfunction, as there are fewer components to break down. [13]
Watts continues:
The bone structures of land animals have many similarities. Does this prove they are all related by a common origin in evolution? Or does it reflect a deeper underlying imperative, determined, for example, by the materials available, so that what we observe represents the best way to form a skeleton?
Watts does not mention that aquatic vertebrates such as whales and dolphins also share many similarities with terrestrial vertebrates despite the fact that they inhabit radically different environments. The underlying similarity in skeletal anatomy in tetrapods is not proof of common descent, but it is entirely consistent with what one would expect if all tetrapods shared a common ancestor, with the anatomical differences arising from a process of descent with modification.
The mammalian forelimb has many different functions depending on the species being examined. In bats, it serves as a wing. In dolphins, it is a paddle. Monkeys use theirs for grasping while horses use theirs for locomotion. Moles use their forelimb for digging. Despite the fact that each use is radically different, the underlying structure of these limbs is the same. We would expect this with common descent, but a designer is not compelled to reuse the same underlying structure, but is free to design the best possible upper limb.
 |
| Source: Wikipedia |
One of the most powerful arguments for common descent are shared quirks of anatomy. One notable example is the recurrent laryngeal nerve. This is a branch of the vagus nerve, one of the 12 cranial nerves - so called because instead of branching from the spinal cord, they branch from the brain stem, and exit via other foramina. The recurrent laryngeal nerve is a branch of the vagus nerve and is one of the nerves that innervate the larynx. From its origin off the vagus nerve to its destination at the larynx is a distance of only a few centimetres. However, instead of passing directly to the larynx, it takes a totally pointless detour down the neck into the chest, loops under the aorta (the largest artery in the body, which emerges directly from the left ventricle of the heart) and then ascends up the neck to the larynx. There is no reason for this detour.
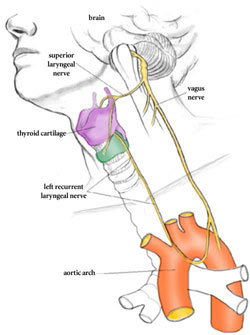
The giraffe is the most spectacular demonstration of the path of the recurrent laryngeal nerve. As with other mammals, it goes past the larynx, down its long neck, under its aorta and up to the larynx:
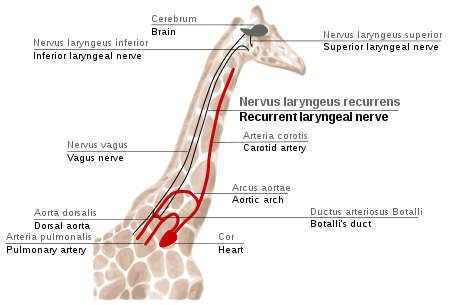 |
| Source: Wikipedia |
The evolutionary biologist Mark Ridley explains the reason behind this anatomical quirk:
Evolution by natural selection proceeds in small, local steps and each change has to be advantageous in the short term. Unlike a human designer, natural selection cannot favour disadvantageous changes in the knowledge that they will ultimately work out for the best. As Wright emphasised in his shifting balance model...natural selection may climb to a local optimum, where the population may be trapped because no small change is advantageous, though a large change could be. As we saw, selection itself (when considered in a fully multidimensional context), or neutral drift, may lead the population away from local peaks; but it also may no. Some natural populations now may be imperfectly adapted because the accidents of history pointed their ancestors in what would later become the wrong direction.
The recurrent laryngeal nerve provides an amazing example. The laryngeal nerve is, anatomically, the fourth vagus nerve, one of the cranial nerves. These nerves first evolved in fish-like ancestors. As figure 10.12a shows, successive branches of the vagus nerve pass, in fish, behind the successive arterial arches that run through the gills. Each nerve takes a direct route from the brain to the gills. During evolution, the gill arches have been transformed; the sixth gill arch has evolved in mammals into the ductus arteriosus, which is anatomically near to the heart. The recurrent laryngeal nerve still follows the route behind the (now highly modified) "gill arch": in a modern mammal, therefore, the nerve passes from the brain, down the neck, round the dorsal aorta, and back up the larynx.
In humans, the detour looks absurd, but it is only a distance of a foot or two. In modern giraffes, the nerve makes the same detour, but it passes all the way down and up the full neck of the giraffe's neck. The detour is almost certainly unnecessary and probably imposes a cost on the giraffe (because it has to grow more nerve than necessary and signals sent down the nerve will take more time and energy). Ancestrally, the direct route for the nerve was to pass posterior to the aorta; but as the neck lengthened in the giraffe's evolutionary lineage the nerve was led on a detour of increasing absurdity. If a mutant arose in which the nerve went directly from brain to larynx, it would probably be favoured (though the mutation would be unlikely if it required a major embryological reorganization); the imperfection persists because such a mutation has not arisen (or it arose and was lost by chance). The fault arose because natural selection operates in the short term, with each step taking place as a modification of what is already present. This process can easily lead to imperfections due to historical constraint - though most will not be as dramatic as the giraffe's recurrent laryngeal nerve. [14]
 |
| Evolution of the recurrent laryngeal nerve. (a.) In fish, the vagus nerve sends direct branches between successive gill arches. (b.) In mammals, the gill arches have evolved into a very different circulatory system. The descendant nerve of the fish's fourth vagus now passes from the brain, down to the heart (in the thorax) and back up to the larynx. Redrawn, by permission of the publisher, from Strickberger (1990), modified from de Beer (1971). |
Watts attempts to rebut the evidence for suboptimal design. He argued:
Physiotherapists and others are always telling us that the human spine is badly designed, but can they explain how to make it better and still perform all its everyday functions.
Special creationists invariably invoke 'design' and 'beauty' in nature as 'evidence' of special creation, but are curiously silent when demonstrably flawed design such as the human vertebral column is highlighted. Evolutionary biologists are not obliged to show how the spine should have been designed in order to eliminate scoliosis, vertebral fracture and disc herniation. What they have shown is that the human spine - as one would expect if common descent was true and we are descended from quadrupeds - is based on a quadruped design, and is susceptible to injury unique to humans:
However, in turning a spine originally adapted for a quadruped into one that is perpendicular to the ground has resulted in numerous problems that are unique to our species. Included in such uniquely human maladies are spondylolysis, spondylolosthesis, herniated discs, spontaneously fractured vertebrae, kyphosis, and scoliosis. An evolutionary perspective sheds light on why we remain uniquely vulnerable to such a wide range of spinal disorders. [15]
In fact, anatomists and developmental biologists have been pointing out how the human body's quirks these make sense only in the light of an evolutionary origin of the human body. [16]
Despite the fact that humans are not hatched from eggs, we still make a vestigial yolk sac. Occasionally it persists as a Meckel’s diverticulum of the ileum that can lead to an umbilical fistula which can result in inflammation, perforation and bleeding. It is a developmental legacy inherited from reptile-like ancestors that did lay eggs.
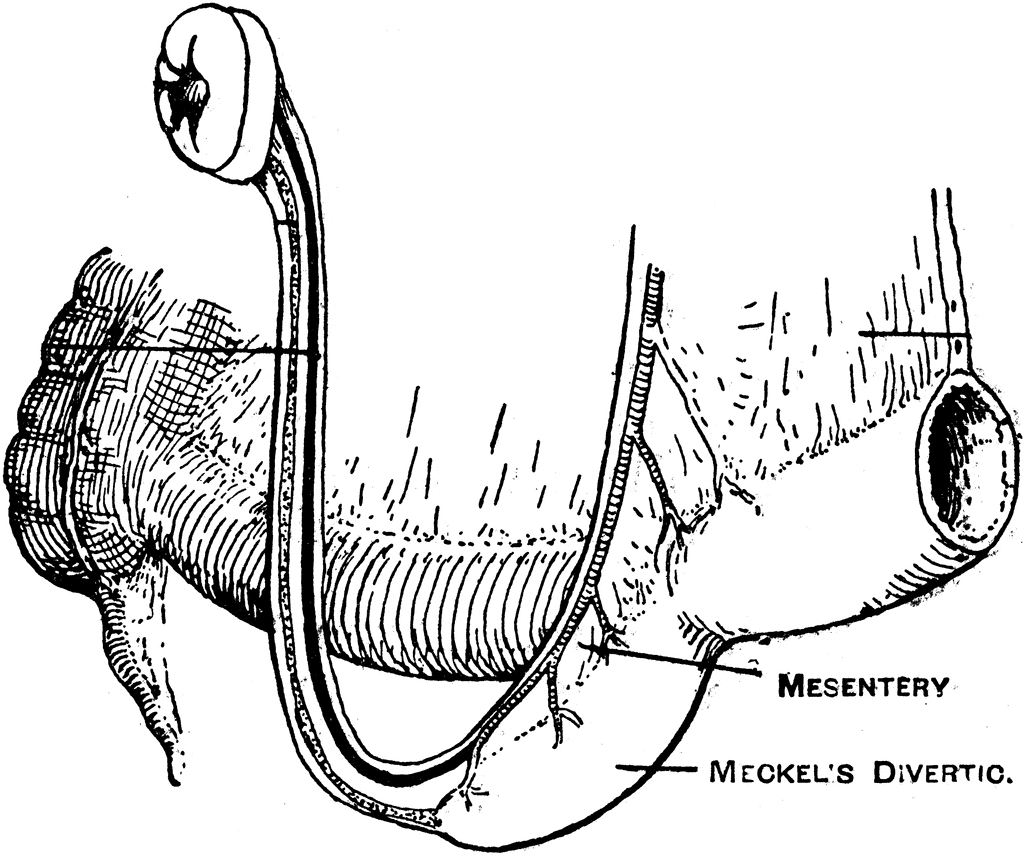
Meckel's diverticulum, the remains of he vitello-intestinal duct, and of the artery to the yolk sac.
Source: Treves, Frederick Surgical Applied Anatomy (New York: Cassell and Company, LTD, 1922) 412
In addition to a vestigial yolk sac, we also have the remnant of the vitellogenin gene which in reptiles codes for a protein component of egg yolk. The human vitellogenin pseudogene is in exactly the same place in the genome as it is in reptiles, a finding which makes sense only in the light of evolution. [17] One could go on, but the point has been made.
As Watts continues, he diverges considerably from the initial question of whether similarity in genetic sequences is evidence for evolution, touching briefly on the apparent constraints made on life by the properties of carbon and water and the alleged improbability of abiogenesis. None of this has any relevance to the original question which has essentially been ignored.
Watts continues with a brief examination of enzymes, which catalyse chemical reactions so they proceed at a much faster rate than they normally would go. He says:
These enzymes have ‘active centres’ that accomplish this remarkable feat; but in order to have the requisite activity, these centres are very restricted in their chemical structure. Thus an enzyme in a snail performing the same reaction found in a dog will almost certainly have a very similar active centre, and we may suspect that the differences are more to do with optimising the performance of the molecule for activity in snail and dog than with chance.
Watts has missed the point completely. Coding redundancy means there are many ways in which a protein of a given length can be encoded. As we saw earlier, for cytochrome c, there are approximately 3^104 or 4.1 x 10^49 possible ways to encode for the same 104 amino acid sequence. There is no reason to favour one possible coding sequence over another, but in fact when we look at the coding sequences for cytochrome C in life, we find them clustering together in a way consistent with common descent. Any argument that each species needs its particular unique cytochrome C is undermined by the fact that human [18] and rat [19] cytochrome C have been inserted into yeast cells and functioned adequately.
His argument that enzymes are very restricted in their chemical structure is not always true. Again, with respect to cytochrome c, we now that only 33% of the amino acids are necessary for it to function. In other words, most of the amino acids can be replaced by functionally similar amino acids. This means that a functional cytochrome C enzyme does not need to be specified by the same 104 amino acids. Given the fact that we now have amino acid as well as coding redundancy, the potential coding space for a functioning cytochrome c enzyme is considerably larger than 3^104 possible genes.[20] The number has been calculated [21] as being around 2x10^93. Despite this unimaginably large potential coding space, the cytochrome c sequences of humans and chimpanzees are identical. Giving humans and chimpanzees completely different cytochrome C sequences would have been trivial for an intelligent designer, and would have been difficult to reconcile with common descent. One is entirely justified in the face of this evidence to conclude that common descent is the most reasonable explanation for the molecular genetics evidence.
Watts concludes:
In sum, then, within the world of living things all closely dependent on one another, using the same basic building blocks and chemistry, it is hardly surprising to see similarities as well as great variations.
This is yet another error Watts made in his article. Mainstream scientists do not cite mere similarities in biochemistry as proof of common descent. Trivially, if common descent was true, we’d expect to see all organisms using essentially the same genetic code. Common descent is well attested by gene homology, synteny and shared ‘genetic errors’ none of which can be readily explained by special creation. In fact, just the evidence from shared endogenous retroviral elements between humans and apes alone is enough to make the case clear.
Again, while one takes no delight in criticising an article by a fellow believer, we need to be honest with the evidence, no matter where it leads. Watts has made sweeping claims in a number of fields ranging from comparative genomics to palaeontology in which he is clearly abysmally informed. To say that this reflects poorly on us is an understatement.
References
1. Watts J "Evolution or Creation? The argument from comparative anatomy." The Testimony January 2005 p 31-33
2. Gregory TR “Evolution as Fact, Theory and Path” Evo Edu Outreach (2008) 1:46-52
3. ibid p 47
4. Gould, Stephen Jay. “Evolution as Fact and Theory” in Hens Teeth and Horse's Toes: Further Reflections in Natural History. (1983 New York: W. W. Norton & Co.) p 260.
5. Thewissen JGM, Cooper LN, George JC, Bajpai S. From Land to Water: the Origin of Whales, Dolphins, and Porpoises. Evolution: Education and Outreach. 2009;2(2):272-288.
6. Prothero DR. Evolutionary Transitions in the Fossil Record of Terrestrial Hoofed Mammals. Evolution: Education and Outreach. 2009;2(2):289-302.
7. Clack JA. The Fish–Tetrapod Transition: New Fossils and Interpretations. Evolution: Education and Outreach. 2009;2(2):213-223.
8. Chiappe LM. Downsized Dinosaurs: The Evolutionary Transition to Modern Birds. Evolution: Education and Outreach. 2009;2(2):248-256.
9. Prothero D.R. “Evolution: what the fossils say and why it matters” (2007 Columbia University Press) p 134
10. ibid, p 179-183
11. Hedges S.B. Kumar S “The Timetree of Life” (2009, Oxford University Press)
12. Novella S “Suboptimal Optics: Vision Problems as Scars of Evolutionary History” Evo Edu Outreach (2008) 1:493–497
13. ibid, p 496
14. Ridley M "Evolution" (2004, Wiley-Blackwell) p 281-2
15. Latimer B "A Backache of Longstanding: An Evolutionary Perspective on the Human Vertebral Column" Abstract of talk presented at the 2013 American Association for the Advancement of Science Meeting.
15. Latimer B "A Backache of Longstanding: An Evolutionary Perspective on the Human Vertebral Column" Abstract of talk presented at the 2013 American Association for the Advancement of Science Meeting.
16. Held L.I. “Quirks of Human Anatomy” (2009, Cambridge University Press)
17. Venema DR “Genesis and the Genome: Genomics Evidence for Human-Ape Common Ancestry and Ancestral Hominid Population Sizes” Perspectives on Science and Christian Faith (2010) 62:166-178
18. Tanaka, Y., Ashikari, T., Shibano, Y., Amachi, T., Yoshizumi, H., and Matsubara, H. (1988) "Construction of a human cytochrome c gene and its functional expression in Saccharomyces cerevisiae." J Biochem (Tokyo) 103: 954-61
19. Scarpulla, R. C., and Nye, S. H. "Functional expression of rat cytochrome c in Saccharomyces cerevisiae." Proc Natl Acad Sci (1986) 83: 6352-6
20. Dickerson, R. E., and Timkovich, R. cytochrome c. The Enzymes. (1975 P. D. Boyer. New York, Academic Press). 11: 397-547.
21. Yockey, H. P. (1992) Information Theory and Molecular Biology. (1992 Cambridge University Press) p 254


Comments
Post a Comment