Why Wrested Scriptures is wrong on evolution - Part 3
The last post looked at Ron Abel’s
failure to properly define evolution by referring to terms such as ‘gradual
change in the characteristics of species over time’, ‘horizontal
differentiation’ and ‘vertical differentiation.’ This failure to properly
differentiate between evolution as fact and evolution as theory meant that Abel’s
attack on evolution was fundamentally flawed.
Abel’s failure to properly define
evolution can be seen in his attempted rebuttal of the evidence for ‘vertical
evolution’, which bears some resemblance to the fact of common descent and
large-scale evolutionary change. Abel recognised four of the five main lines of
evidence for evolution:
2. It is useful to anticipate that the evidence for 'vertical evolution' is usually taken from four major areas:
- Paleontology - the study of fossil remains of extinct animals and plants, including traces of their existence (e.g. footprints in slate, clay, or coal). It attempts to establish the order in which things lived.
- Morphology and Comparative Anatomy - the study of the comparison of living and dead things for resemblances and differences which might suggest relationships between them and indicate whether one form might have been derived from another.
- Embryology - the study of developing creatures before their birth. Comparisons are made between the adult or developing forms of other creatures.
- Genetics - the study of how the characteristics of parents are transmitted to their offspring, and how variations in these characteristics can arise. It is also concerned experimentally with breeding research in which new variables are produced and specialized.
Missing however is biogeography,
which is alluded to only briefly in a footnote. This oversight is damning as
the biogeographical distribution of species is one of the strongest lines of
evidence for common descent, and shows how rudimentary Abel’s understanding of
evolution was. Furthermore, his definition of the evidence for evolution from
genetics is not referring to the evidence for common descent from comparative
genetics, but rather to the theoretical mechanism behind evolutionary change. This
conflation of evolution as fact with evolution as theory stems directly from
his flawed definition of evolution.
Before looking at the evidence
for common descent from palaeontology, genetics, comparative anatomy,
embryology and biogeography, a comment on one of Abel’s footnotes would not go
astray. Abel claimed that the main evidence for ‘vertical evolution’ came from
four areas, and stated in a footnote that:
Evidence for evolution is sometimes taken
from biogeography (the study of geographical distribution of plants and
animals) and taxonomy (the science of the classification of organisms). These,
however, are not usually the crucial arguments. When arguments of
classification are advanced by evolutionists it requires stressing that it is
the evolutionist, and not the
fossils, which orders the single celled amoeba, first, and next to it places a
multicellular hydra, a three-layered worm, an amphioxus, and then a fish, an
amphibian, reptile, lower mammal, lower ape, higher ape, and man. The order is
a synthetic one. It no more proves that one form of life came from another than
the arrangement of books on a shelf from simple to complex proves that the
books "evolved". Both simple and complex may have been produced
simultaneously. Arguments from the classification assume the very thing which must be proved.
This is nonsense. As I said
earlier, the biogeographical distribution of species is one of the strongest
lines of evidence for common descent. (More on this later). Furthermore, his
assertion that the classification of organisms is entirely arbitrary is false,
and betrays a complete ignorance of the principles behind taxonomy.
Take a look at the standard
family tree used to classify organisms:
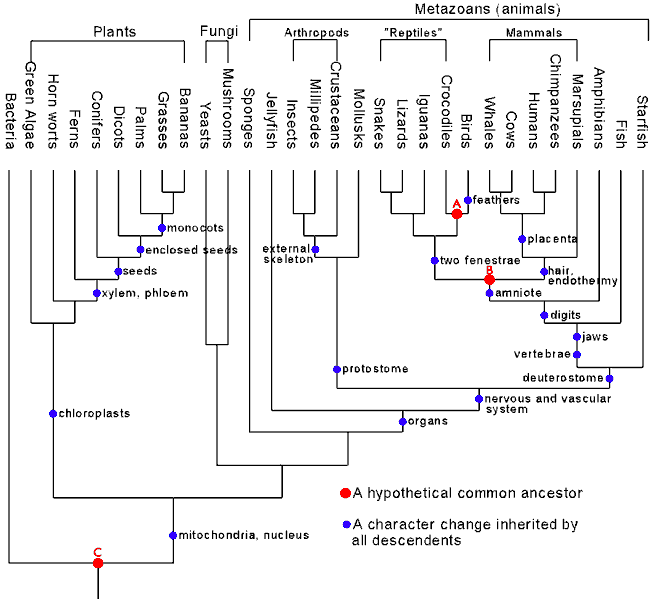 |
| Source: http://www.talkorigins.org/faqs/comdesc/images/phylo.gif |
This classification system is not
arbitrary, but rather consists of a nested hierarchy, that is, groups within
groups. Take mammals, the group in which we are classified. Mammals are defined
by a number of key features such as a lower jaw consisting of a single bone,
three ear bones, and the ability to produce milk. In turn, mammals, birds and
reptiles nest in a large group known as amniotes, which are defined as a group
of tetrapods that have an egg adapted to land life rather than aquatic life. Most
mammals do not lay eggs, but the anatomical structures in the amniote egg have
their equivalents in the placenta.
Amniotes, fish and amphibians in
turn nest within an even larger group known as vertebrates which are defined as
animals that have a vertebral column with a spinal cord dorsal to it, and a gut
ventral to it.
Vertebrates, echinoderms
(starfish, sea urchins and sea cucumbers) and two other smaller groups of
animals themselves fall into an even larger group known as deuterostomes, so
called because they share a common feature of embryonic development where the first
opening in the early embryo eventually becomes the anus.
Deuterostomes and protostomes
(another large group including molluscs and arthropods which are defined by the
fact that the first opening in the early embryo becomes the mouth) together
form an even higher group called bilaterians which are defined by their
possession of bilateral symmetry (clearly defined top, bottom, front and back)
as well as in most cases three distinct layers of cells in their embryos.
Bilaterians, comb jellies,
sponges and jellyfish in turn nest within the metazoans, or animals which are
united by being multicellular, needing to digest food to obtain energy, and
lacking rigid cell walls.
The important thing to stress
about this family tree is that it is not arbitrary, but driven by objective
taxonomic features. Abel was completely wrong when he asserted:
When arguments of classification are
advanced by evolutionists it requires stressing that it is the evolutionist, and not the fossils,
which orders the single celled amoeba, first, and next to it places a
multicellular hydra, a three-layered worm, an amphioxus, and then a fish, an
amphibian, reptile, lower mammal, lower ape, higher ape, and man. The order is
a synthetic one.
As we can see, this nested
hierarchical structure is an objective feature, and not one forced on nature by
scientists trying to prove evolution. Furthermore, the only known process that
generates a nested hierarchy is descent with modification. This leads to a
point overlooked by special creationists. Common descent could easily be
falsified by the discovery of many animals that could not be classified in the
existing nested hierarchical family tree. Every day, common descent is being
tested with the discovery of new animals and plants. Common descent predicts that
they will fit within the existing pattern, and to date, every animal and plant
discovered fits within this predicted tree.
Conversely, if special creation
was true, then there is no reason why animals could be created with features
from different nested hierarchies. Feathers make excellent insulation, so there
is no reason why the Creator could not have given some mammals feathers. Likewise,
given that milk is a good source of food, birds could have been created with
the ability to lactate. Likewise, given that whales lactate, fish could have
been created with mammary glands. But we don’t see this. Everything in nature
falls within a nested hierarchy, exactly as predicted by common descent.
Abel’s complete failure to grasp
the reason behind taxonomy betrayed a fundamental ignorance of the subject he
set out to criticise. It isn’t a trivial mistake. Animals and plants can be
objectively classified in a nested hierarchy, a pattern of groups within groups
which is what descent with modification would produce. This family tree is both an objective method of classification, as well as a prediction of the sort of life we should, and should not expect to find.
A final comment would not go astray here. It is also possible to create a family tree based on molecular sequences or gene sequences. Nearly fifty years ago, Linus Pauling and Emile Zuckerkandl predicted that one of the best available proofs of common descent would be the consonance between family trees from morphological and molecular data:
"It will be determined to what extent the phylogenetic tree, as derived from molecular data in complete independence from the results of organismal biology, coincides with the phylogenetic tree constructed on the basis of organismal biology. If the two phylogenetic trees are mostly in agreement with respect to the topology of branching, the best available single proof of the reality of macro-evolution would be furnished. Indeed, only the theory of evolution, combined with the realization that events at any supramolecular level are consistent with molecular events, could reasonably account for such a congruence between lines of evidence obtained independently, namely amino acid sequences of homologous polypeptide chains on the one hand, and the finds of organismal taxonomy and paleontology on the other hand. Besides offering an intellectual satisfaction to some, the advertising of such evidence would of course amount to beating a dead horse. Some beating of dead horses may be ethical, when here and there they display unexpected twitches that look like life."[1]
When the data from molecular and gene sequencing started appearing, the prediction of Pauling and Zuckerkandl was confirmed as phylogenetic trees from molecular data agreed overwhelmingly with the existing phylogenetic tree. This is exactly what common descent would predict.
Conversely, there is no reason for this to be true if special creation was in fact true. Due to redundancy in the genetic code, there are approximately three different ways to code for each amino acid (the building blocks of proteins). This means that for even a moderately large protein such as cytochrome c which has around 104 amino acids, there are 3^104 or 4x10^49 (that's 4 with 49 zeros after it!) possible ways to code for exactly the same protein. What we see is that the coding sequences for cytochrome c cluster together closely in such a way as produce, when graphed, a family tree remarkably consonant with the existing evolutionary family tree.
 |
| Source: Mulligan, P.K. (2008) Proteins, evolution of in AccessScience, ©McGraw-Hill Companies. |
It is hard to ask for a better demonstration of common descent.
References
1 Zuckerkandl E, Pauling L (1965) "Evolutionary Divergence and Convergence in Proteins." in Evolving Genes and Proteins, p. 101.
[2] Common descent would predict that the gene sequences for all animals which have cytochrome c should cluster together in a pattern consistent with descent with modification. Conversely, if special creation was true, there is an unimaginably large coding space for this protein, enough for each animal to have a unique coding sequence separated by so much space as to falsify irrefutably common descent. We do not see this. Either God has been deliberately creating life in such a way as to simulate common descent down to the molecular level (and for no objective reason given that each of those possible 4x10^49 coding sequences produces exactly the same protein), or the remarkable consonance between molecular and morphological phylogenetic trees should be seen as what Zuckerkandl and Pauling stated, the best available proof of common descent.


Comments
Post a Comment