Why Wrested Scriptures is wrong on evolution - Part 5
Abel’s second attack on what he
called ‘vertical evolution’ once again suffered from his failure to properly
differentiate between common descent and the theoretical mechanism postulated
to explain it. The ‘common designer’ argument fails to take into account the
fact that the ‘common designer’ appears constrained to create in a nested
hierarchical pattern (groups within groups) which is exactly the process one
would expect to see from a process of descent with modification. The fact that
the same anatomical mistakes (inverted retina, recurrent laryngeal nerve) are
found in groups is consistent with evolution, but makes the common designer
look like a serial bungler, repeatedly making the same mistakes in such a way
as to simulate common descent. The consonance between molecular and
morphological family trees is exactly what one would expect from common
descent, but is inexplicable from a special creationist point of view.
His suggested strategy to rebut
the morphological, anatomical and embryological data is highlighted below:
Morphological, Anatomical, and Embryological
- Evidence required - It must be shown that x and y are part of an historical sequence of progressive changes, or that x can be made to give birth to y by this or that genetic modification.
- Evidence lacking
1. Comparing the anatomy of the wing of a bird,
the paddle of a whale and the arm of a man, certain similarities of structure
can be observed. But resemblances of structure are just as capable of
indicating a common designer as ancestry from prior forms.
2. Morphological, anatomical and embryological
evidence depend on palaeontological and genetic claims. It depends on whether
the fossil record shows a finely graded progression from simple to complex, and
whether the study of genetics provides the mechanisms by which creatures would
move over time from simple to complex.
I’ve shown that the morphological
data does indeed support common descent in an earlier post. Once again, life
can be classified objectively in a nested hierarchy, a pattern which is exactly
what descent with modification predicts. Asserting that this is evidence of
common design is effectively saying that God designed life to look as if it had
come about by a process of descent with modification – it is an ad hoc argument
employed to argue away evidence which has convinced > 99% of professional
biologists that common descent is the better way to explain what we see.
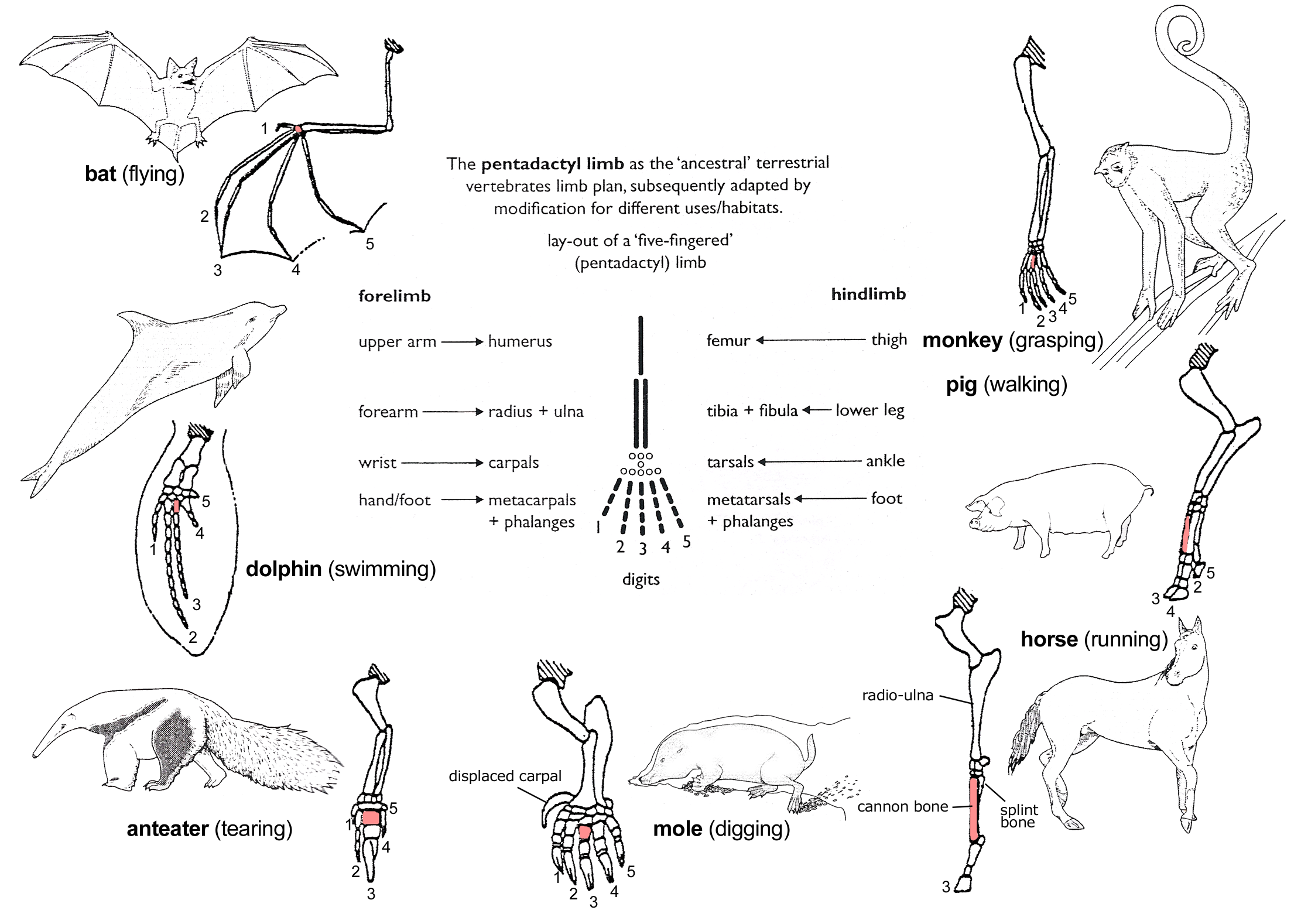
The vertebrate limb is one of the classic examples of how comparative anatomy provides support for common descent. Vertebrates share a common ancestor if common descent is true. Therefore, we'd expect a process of descent with modification to have altered the ancestral vertebrate limb to perform different functions. This is exactly what we see, as the above diagram shows.
Conversely, the common design argument suffers from a number of serious problems:

A knowledge of comparative anatomy and embryology sheds light on this anatomical quirk. In fish, the successive branches of the vagus nerve pass behind the arterial arches running through the gill. Fish do not have a long neck, so this pathway is direct.
Vertebrate embryos initially share this anatomical configuration, which during development is modified. As a result, the path of the recurrent laryngeal nerves loops under arteries such as the right subclavian and the aorta for the right and left RLN, respectively.

A prediction of common descent would be that irrespective of neck length, the RLN would still bypass the larynx, arch under the aorta, and loop up to the larynx. From a design point of view, this would be wasteful and pointless. The extreme case would be that of the giraffe, which would have a couple of metres of wasted nerve. This is exactly what we see:

The vertebrate eye is more evidence for common descent, as it too is an example of shared suboptimal design In the vertebrate eye, the light sensing cells in the retina point away from light. This means light has to pass through blood vessels before reaching the light sensing cells There is a blind spot in vertebrate retinas where blood vessels and nerve fibres pass as they exit and enter the eye. This blind spot would not be needed if the light cells pointed towards light.

Conversely, the common design argument suffers from a number of serious problems:
- There is more than one way to make a wing, grasping limb, flipper, digging limb or running limb, yet we see nothing but variations on the same theme.
- As shown earlier, a comparative anatomical study of life shows nested hierarchies, which means that any designer would be creating in such a way as to simulate common descent. The 'common design' argument then comes across more as as an untestable way to explain away the data
- Shared sub-optimal design is something we'd expect with a blind process such as evolution where initial design errors are inherited by descendant species. An intelligent designer not only can start from a clean sheet, but would not constantly make the same mistakes in the same group.
A knowledge of comparative anatomy and embryology sheds light on this anatomical quirk. In fish, the successive branches of the vagus nerve pass behind the arterial arches running through the gill. Fish do not have a long neck, so this pathway is direct.
Vertebrate embryos initially share this anatomical configuration, which during development is modified. As a result, the path of the recurrent laryngeal nerves loops under arteries such as the right subclavian and the aorta for the right and left RLN, respectively.
A prediction of common descent would be that irrespective of neck length, the RLN would still bypass the larynx, arch under the aorta, and loop up to the larynx. From a design point of view, this would be wasteful and pointless. The extreme case would be that of the giraffe, which would have a couple of metres of wasted nerve. This is exactly what we see:
This results in an increased susceptibility towards eye disease. Neurologist Steve Novella notes:
Even worse than the backward arrangement of the rods and cones is the fact that the blood vessels that feed the retinal sit on top of the retina — between the light source and the receptive layer. A more logical arrangement would be to have the blood vessels feed the retina from behind, so that they do not get in the way. In healthy eyes, the blood vessels do not cause any perceptible problem (but they are also partly responsible for the blind spot), but they do limit the total amount of light reaching the rods and cones. The real problem is that they are vulnerable to various diseases. [1]Retinal detachment is one complication of having an inverted retina. If the retina was not inverted, the nerve fibres from the photoreceptors would anchor them securely to the layers beneath them. Other problems arising from having an inverted retina stem from having blood vessels on top of the photoreceptor layer, which means that abnormal blood vessel growth (diabetic retinopathy), bleeding (retinal haemorrhage) or inflammatory changes result in reduced vision.
Strangely enough, animals exist with a 'proper' retinal design, where the photoreceptors are in front of the nerve fibres and blood vessels and point towards the light. They are the cephalopods:

The cephalopod eye design is more sensible as the light cells point towards the light, with blood vessels and nerve fibres behind the light sensing layer. The retina is firmly attached to the eye, reducing the likelihood of retinal detachment considerably. Likewise, macular degeneration, and diabetic retinopathy would not be a problem if our eye employed this design. From a special creationist perspective, the vertebrate eye is inexplicable as it is markedly inferior to the cephalopod eye. From the perspective of our evolutionary history, it makes perfect sense.
Embryology
Developmental biologists have recognised for quite some time that early in their development, vertebrate embryos resemble each other closely:
- All have a notochord and dorsal nerve cord
- All have pharyngeal arches
- All have bodies made from repeated muscle blocks or somites
- All have tails

This is known as the phylotypic stage, and poses difficult questions for special creationists. One of them focuses on the shared, counterintuitive way in which the vertebrate face is formed. Structures in the embryonic vertebral arches eventually become facial features.


In all vertebrates, arches 1 and 2 always form the mandible and hyoid bone, respectively. The 3rd and subsequent arches form different structures:
- Fish: the gills
- Humans: throat cartilage, as well as sundry other muscles and glands
A striking example of how embryology provides powerful evidence for common descent comes from the location of the anatomical structures involved in calcium metabolism. (The argument is greatly indebted to P.Z. Myers' post here.) Gills in fish are not only responsible for gas exchange, but in regulation of electrolyte balance. The gills contain sensors and ion pumps that regulate serum calcium levels, making sure that the levels remain at an optimum level.

In humans, we don't have gills, but rather have glands in the neck - the parathyroids and thyroid - to regulate calcium levels. If calcium levels drop too low, then the parathyroid glands signal the kidneys to retain calcium and the bones to release calcium into the blood. Conversely, high calcium levels signal the thyroid which in turn reduces calcium release from bones.
Now, if common descent was true, we would hypothesise that the calcium regulating areas in fish and humans should have the same embryonic origin, as they would have inherited this from the remote common ancestor of fish and humans. We know that in fish, calcium regulation takes place in the gills, which have their embryonic origin in the third and subsequent pharyngeal arches. In humans, the third and subsequent pharyngeal arches form structures in the neck, which is where we find the thyroid and parathyroid glands. I need to stress that there is absolutely no need for these glands to be in the neck. In fact, patients who have their entire thyroids surgically removed due to disease will have their parathyroid glands reimplanted in other anatomical locations.

Impressive as this is, one can go further. In order to confirm whether the parathyroid glands have their embryonic origin in the pharyngeal arches, just as the fish gill sensors have their origin in the pharyngeal arches, we can use molecular biology to test their origin. A molecular marker unique to parathyroids is found in the 3rd and 4th pharyngeal arches in tetrapods. The same marker is found in the pharyngeal arches of fish, even though they don’t have parathyroid glands.
We can test this idea by creating fish that lack the ability to synthesise this marker. These genetically modified fish embryos fail to grow gills. This evidence from anatomy, physiology, molecular biology and developmental biology provide strong evidence supporting the common ancestry of modern fish and modern tetrapods. The common design argument falls apart, as one is entitled to ask why God creates vertebrates in such a way as to consistently simulate common descent.
An earful of jaw - the evolution of the mammalian ear
Perhaps the most spectacular demonstration of common descent from embryology is the developmental biology of the ear and jaw in reptiles and mammals. Mammals have three middle ear bones and one jaw bone, while reptiles have more than one bone in their jaw but only a single middle ear bone.

As both are believed to have a common ancestor, common descent would predict that two bones in the common ancestor which in reptiles form the jaw would in mammals move to form the middle ear. This is exactly what we see in reptile and mammal embryology. The same two embryonic structures that in reptiles form part of the jaw form two of the middle ear bones in mammals.
When we look at the embryology of the human ear in detail, we see:
- Meckel’s cartilage in the embryo is the cartilaginous bar of the mandibular arch.
- The dorsal ends ossify to form the incus and malleus, while the ventral ends ossify at the tip of the jaw near the chin.
- The middle piece of the cartilage vanishes

This developmental pathway makes sense in the light of common descent and the ear’s evolutionary origin. Once again, the special creationist argument is to simply claim 'common design', without bothering to explain why this 'design' constantly mimics what we'd as a result of common descent. Parsimony favours common descent.
Vestigial structures and atavisms
Vestigial traits are structures which served a function in the ancestor of a species, but either have lost their original function, or have been recruited for other uses. They abound in nature, and provide considerable evidence for common descent. Examples include the wings of flightless birds and the remnants of eyes in marsupial moles. Vestigial traits are also completely misunderstood by special creationists who confuse vestigial with useless. This is not the case. Jerry Coyne comments how:
Opponents of evolution always raise the same argument when vestigial traits are cited as evidence for evolution. “The features are not useless,” they say. “They are either useful for something, or we haven’t yet discovered what they’re for.” They claim, in other words, that a trait can’t be vestigial if it still has a function, or a function yet to be found.
But this rejoinder misses the point. Evolutionary theory doesn’t say that vestigial characters have no function. A trait can be vestigial and functional at the same time. It is vestigial not because it’s functionless, but because it no longer performs the function for which it evolved. The wings of an ostrich are useful, but that doesn’t mean that they tell us nothing about evolution. Wouldn’t it be odd if a creator helped an ostrich balance itself by giving it appendages that just happen to look exactly like reduced wings, and which are constructed in exactly the same way as wings used for flying?
Indeed, we expect that ancestral features will evolve new uses: that’s just what happens when evolution builds new traits from old ones. Darwin himself noted that “an organ rendered, during changed habits of life, useless or injurious for one purpose, might easily be modified and used for another purpose.” [2]Vestigial traits abound in nature; some of the better known examples include:
Dewclaws. These are vestigial digits that can be found on the feet of many vertebrates. In animals that walk on their digits such as dogs and cats, they often do not reach the ground.

Piloerection. This reflex occurs either during cold weather or in response to stress. In the latter, it causes animal to appear larger and more intimidating, while in the former it traps an insulating layer of air. In humans, the response serves neither function and is a relic of our animal ancestry.

Coccyx. Otherwise known as the tail bone. During embryogenesis, human embryos grow a tail, which later degenerates into the coccyx. Its only function now is to serve as an attachment for muscles. The genes for making a tail still exist in humans, and examples of functional tails that contain muscle, nerves and are capable of movement exist.

The top diagram compares the lower spine and tail of a lemur with the lower spine of humans, where the corresponding vertebral bodies C1-C4 (not to be confused with the cervical vertebrae) are fused to form the coccyx. The lower image shows the persistence of the embryonic tail in a baby, and shows the presence of distinct vertebral bodies. (Source: Coyne - Why Evolution is True)
Tooth development in baleen whales. Despite not having teeth, baleen whale embryos grow tooth buds in utero, only to break them down again later in embryogenesis. Transitional fossils of whales with both teeth and baleen-related structures exist, so we know baleen whales evolved from toothed whales. Molecular evidence corroborates this fact as modern baleen whales have broken genes that code for enamel-related protein. The presence of broken genes coding for enamel-related protein in whales that do not have teeth, not to mention the embryonic development of tooth buds that are later broken down is very much an example at the gross anatomical and molecular level of a half-built structure. From an evolutionary point of view, it is intelligible, but from the perspective of special creation, it is inexplicable.

Tooth buds in the jaws of baleen whale embryos
Palmaris longus. in humans, this upper limb tendon which lies between the flexor carpi ulnaris
and the flexor carpi radialis muscles has no appreciable function, and
in fact is absent in between 10-15% of people. It is often
harvested as a tendon graft in hand surgery without any functional
deficit. It functions in other animals to expose claws – in humans, this
function is no longer necessary and this muscle has become vestigial.
Plantaris. this lower limb muscle has minimal functionality and is absent in up to 10% of the population. As with the palmaris longus, it is often harvested for use in reconstruction surgery with no marked functional deficit, as plantarflexion of the foot is carried out mainly by the gastrocnemius and soleus muscles. In primates, it is used for grasping with the feet, but as we are an obligate bipedal species, we have lost the ability to grasp with the feet and no longer need this function. Again, this is very much an example of a vestigial, “half-functional” structure whose existence makes sense in the light of evolution.
Apomixis in flowering plants. this refers to asexual reproduction via seeds, and is found in a number of plant genera including the dandelions. Put simply, these plants have flowers, despite the fact they do not need them for reproduction. Flowers in apomictic plants are very much a vestigial structure.
Toenails in manatees. these aquatic mammals have flippers and no separate digits, but still retain toenails.

Flightless beetles. there are many flightless beetle species such as the Kauai Flightless Stag Beetle which have fused outer wing covers, but still have inner wings present in a vestigial state. Recognition of their evolution from beetles that could fly however makes this design flaw understandable.

Atavisms, or 'throwbacks' to use a vernacular term are the presence of ancestral traits in organisms that are not normally expressed. Examples include hind limbs on whales, toes on horses and tails in humans. Their presence is readily explained by common descent. Normally inactive developmental pathways for traits present in ancestral species but not normally expressed in their descendants are for some reason activated, resulting in the appearance of these 'throwbacks':
Plantaris. this lower limb muscle has minimal functionality and is absent in up to 10% of the population. As with the palmaris longus, it is often harvested for use in reconstruction surgery with no marked functional deficit, as plantarflexion of the foot is carried out mainly by the gastrocnemius and soleus muscles. In primates, it is used for grasping with the feet, but as we are an obligate bipedal species, we have lost the ability to grasp with the feet and no longer need this function. Again, this is very much an example of a vestigial, “half-functional” structure whose existence makes sense in the light of evolution.
Apomixis in flowering plants. this refers to asexual reproduction via seeds, and is found in a number of plant genera including the dandelions. Put simply, these plants have flowers, despite the fact they do not need them for reproduction. Flowers in apomictic plants are very much a vestigial structure.
Toenails in manatees. these aquatic mammals have flippers and no separate digits, but still retain toenails.
Flightless beetles. there are many flightless beetle species such as the Kauai Flightless Stag Beetle which have fused outer wing covers, but still have inner wings present in a vestigial state. Recognition of their evolution from beetles that could fly however makes this design flaw understandable.
Brachypterous Colophon westwoodi with vestigial wing visible. [image M. Cochrane, Iziko © ]
Atavisms, or 'throwbacks' to use a vernacular term are the presence of ancestral traits in organisms that are not normally expressed. Examples include hind limbs on whales, toes on horses and tails in humans. Their presence is readily explained by common descent. Normally inactive developmental pathways for traits present in ancestral species but not normally expressed in their descendants are for some reason activated, resulting in the appearance of these 'throwbacks':
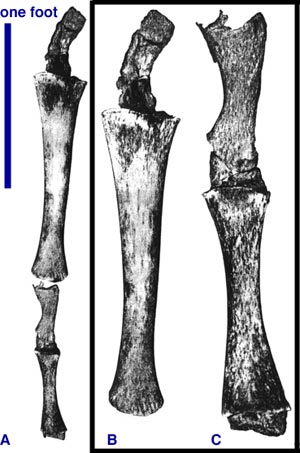
Atavistic hind limb bones in a humpback whale. Source: TalkOrigins Archive

Hind limbs in a dolphin. Source: TalkOrigins Archive
This is hardly surprising given common descent, but inexplicable from special creation. After all, if each animal was specially created, why include the developmental pathways for traits that are simply not needed, and more importantly, exactly the pattern of traits which would be expected if common descent were true. Eventually, the invocation of 'common design' ceased to have any explanatory power and comes across as simply a desperate attempt to explain away an avalanche of evidence for common descent.
References
References
1. Novella S "Suboptimal Optics: Vision Problems as Scars of Evolutionary History" Evo Edu Outreach (2008) 1:493-497
2. Coyne, J, Why Evolution Is True, (2009: OUP) p 62.
2. Coyne, J, Why Evolution Is True, (2009: OUP) p 62.

