"20 scientific facts seldom taught to students" critically reviewed #18
Collyer's eighteenth 'scientific fact' is the age-old special creationist blunder that there are no transitional fossils: "The so-called 'missing link' between one form of life and another requires many millions of missing links of a slow evolutionary process did actually take place. All are missing."
Wrong. Collyer's "20 scientific facts" series is studded with errors, but his assertion that there are no transitional fossils is particularly egregious, given that as renown palaeontologist Stephen Jay Gould pointed out, "[t]ransitional forms are generally lacking at the species level, but they are abundant between larger groups." [1]
The reason why transitional forms are generally lacking at the species level is due partly to the incompleteness of the fossil record, but mainly due to the mechanisms of speciation, in which species develop quickly in a sub-population in a small part of the geographical extent of the ancestral species.
Finally, his reference to 'missing links' betrayed yet again lack of familiarity with the subject. Creationists who refer to 'missing links' are thinking of evolution in terms of a ladder from simple organisms to complex organisms. Evolution however is modelled not by a ladder, but by a low bushy tree. Missing links are missing only because there are no such thing as 'missing links.' Rather, palaeontologists look for fossils with transitional features, and the fossil record abounds with them. [2]
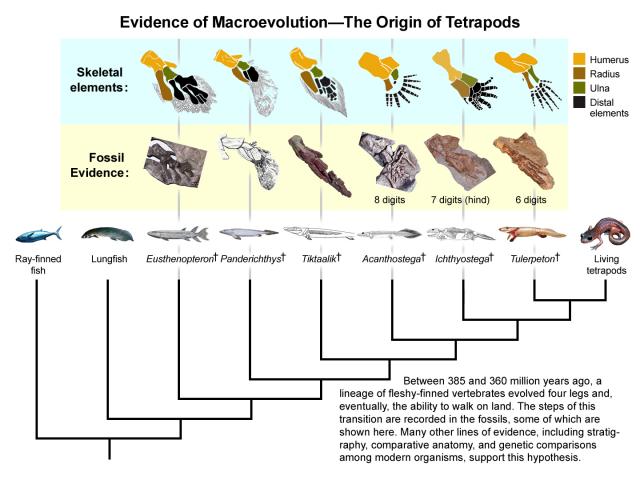
Special creationists such as Collyer who claim that there are no missing links are wrong. The fossil record is replete with transitional fossils. For example, the evolution from lobe finned fish to tetrapod is very well documented, with fossils such as Tiktaalik roseae, a Devonian fish, having a wrist, and other tetrapod features. [3-6] The evolution of whales [7], terrestrial hoofed mammals [8], humans [9], birds (from dinosaurs) [10] and mammals from reptile like tetrapods [11] are all well documented. While recent years have seen an explosion in our understanding of the fossil record, by the second half of the 20th century, the evidence for evolution from the fossil record was robust. As early as 1960, TN George could say that:
There is no need to apologize any longer for the poverty of the fossil record. In some ways it has become almost unmanageably rich, and discovery is out-pacing integration: the growing number of species of Formaminifera that remain undescribed in the cabinets of the oil companies probably is of the order of thousands; and while most other organic groups are not so fully collected the ratio of added finds to palaeontologists studying them is constantly expanding. But what remains to be discovered is likely to be of less and less radical importance in revealing major novelties, more and more of detailed infilling of fossil series whose outlines are known. The main phyla, in so far as they are represented by fossils, now have a long and full history that is made three-dimensional by a repeatedly cladal phylogeny. The gaps are being closed not only by major annectant forms, the "missing links" that Darwin so deplored, like the fish-amphibian ichthyostegids, the amphibian-reptile seymouriamorphs, and the reptile-mammal ictidosaurs, but also by new discoveries of phyletic affiliations, as in graptolite structure...Together, the discovery of new fossil forms, the filling out of the details of bioserial change, the interpretation of biofacies, the adoption of new techniques both in fossil morphology and in fossil manipulation, and the establishment of a progressively refined timescale contribute to a present-day palaeontology offering the strongest support, the demonstrative "proof," of the fact and the process of evolution in terms wholly concordant with the essence of Darwinian theory. [12] (Emphasis mine)
The evolution of the mammalian inner ear from reptilian jaw bones is well established both from embryology (the same bone structures that in reptiles form part of the jaw in mammals form part of the middle ear) and palaeontology. Palaeontologist Kevian Padian, in his trial evidence for Kitzmiller v. Dover points out that fossil evidence for this has been known for decades:
Q. Let's talk about mammals. One of the examples that's referenced in [a creationist textbook] is the mammalian ear, inner ear. Could you talk to us about how [the creationist textbook] discusses the mammalian ear and what science shows about that? And you've prepared a demonstrative for this?
A. I put a couple of slides together about the transition in the evolution of the mammal ear, which is unusual compared to all the other vertebrates. The next slide I think shows a bit about this. This is going to get a little complex anatomically, but I hope it will only hurt for a minute. The bones of the middle ear, mammals have three of them. You might have heard of them as the hammer, the anvil, and the stirrup.
The stirrup is a bone that's always in the ear, but the mammals have this anvil and hammer thing which are just outside that stirrup bone. These anvil and hammer bones actually correspond to bones that previously made up the upper and lower jaw joint in all the other animals, not just reptiles or anything like them, but everybody pretty much. So the [creationist textbook] authors claim that to make this correspondence is really stretching it, because they said there's no fossil record of this amazing process.
Consider, that to make this change one of these bones had to cross the hinge from the lower jaw into the middle ear region of the skull. Again this is from [creationist textbook] page 121. So they're saying there's no record of this process and it would be an amazing thing to have to change. The next slide shows that there are actually many sources going back several decades that differ, and there are just a few of them there.
The first one was actually an article by Romer, who was the dean of American vertebrate paleontology for half the century, about a cynodont that has an incipient mammalian jaw articulation, and I'll show you what that is in a minute. That comes from the journal Science in 1969. Here's a somewhat later paper by Edgar Allen of Madison, and now it's Chicago, on the evolution of the mammalian middle ear, and then a third one I put there is very recent piece, a little piece in Science by Thomas Marin from Germany and Zhe-Xi Luo, who's curator at the Carnegie museum here in Pittsburgh just a few hours away, one of the great museums in the country, and they are talking about the evolution of these bones in the middle ear something that is uncontroversial as a principle in comparative anatomy of vertebrates in palaeontology.
Q. Now, I note that first article I believe was from 1969.
A. Was.
Q. So this isn't a new development?
A. Oh, no. Oh, no. It's been known for decades. [13]

Evolution of mammalian jaw joint (Source)
The 1969 paper by Romer to which Padian refers states in the abstract:
A diagnostic mammalian character is jaw articulation between squamosal and dentary bones, replacing the quadrate-articular joint of reptiles. A newly discovered Argentinian Middle Triassic form shows, for the first time in an ancestral reptile, definite evidence of a squamosal-dentary articulation supplementary to the persistent primitive connection. (Emphasis mine) [14]
In other words, Romer describes a reptile with both reptilian and mammalian jaw articulation. In other words, what we have is a transitional fossil. This was decades before Collyer's book was published, and he would have known it if he'd conducted adequate research. Instead he asserted that there are no missing links despite what George and Romer had stated in the 1960s.
There are no missing links because there is no such thing as a missing link
Collyer erred by using the term “missing link”, despite the fact that palaeontologists do not use the term, but refer to transitional fossils, or transitional features on fossils. This is not mere semantics, but refers to the fact that evolution does not proceed in a ladder from simple to complex, but is better modelled as a bushy tree. Implicit in the use of the term “missing link” is the scala naturae, or Great Chain of Being, which has microbes at the bottom, and man at the top. Humans rather are merely one tip of a branching tree, rather than the pinnacle of evolution, and share common ancestry with all forms of life. Louise Mead, an evolutionary biologist associated with the National Center for Science Education makes this point:
The concept of a “missing link” is an “archaic expression”...tracing back to the Great Chain of Being, a view of the physical and metaphysical world as an unbroken chain. It was later temporalized by the evolutionary thought of the eighteenth and nineteenth century to the idea of evolution as a progressive climb up a ladder...These views of evolution create the false expectation that there should be fossil evidence showing “a complete chain of life from simple to complex”...Creationists rely on such views to support their arguments against macroevolution, in particular by pointing out the “conspicuous” absence of “large numbers of intermediate fossil organisms”...using what is still unknown to question whether evolution has occurred. I will deal with the misconception of evolution as a ladder-like progression shortly, but should the fossil record be expected to reveal all species that have ever lived? Clearly not. Knowledge of the fossil record will never be comprehensive...First, there is too much of the Earth to explore, and paleontologists have to be content with samples. Second, given our knowledge of geology, we understand that not all organisms will be fossilized and that there will be systematic biases in what organisms are fossilized...Therefore, any statements rest on a fallible, if informed, assessment of the necessarily incomplete evidence. [15]

Mead’s point about the inherent biases in the fossilisation process is often overlooked by special creationists. Hard-bodied animals fossilise far more readily than soft-bodied animals. Marine environments are far more likely to allow fossilisation than forests, in which fossilisation rarely occurs. Not only do special creationists attack a straw man when they advance the “missing links are missing” canard, they show specific ignorance of taphonomy, the science behind the decay and fossilisation of dead animals.
To this one can add the fact that palaeontologists and evolutionary biologists do not look for intermediate forms, but rather try to reconstruct evolutionary history using shared derived characteristics, or synapomorphies. Furthermore, special creationists overlook the fact that the two lines diverging from a common ancestor will have changed considerably since the lineages split, so this common ancestor will not look intermediate between the current descendants of those lineages. Mead notes:
Today, evolutionary biologists and paleontologists do not focus on finding “intermediates” but rather on reconstructing evolutionary relationships and history using shared derived characters, or synapomorphies. Willi Hennig revolutionized systematics in the 1960s with the introduction of cladistics, which ushered in a new method of phylogenetic analysis and a new approach to systematics. Instead of relying on a Linnaean system of classification, cladistics placed the focus on evolutionary history, specifically identifying features as ancestral (general) or derived (evolved after the lineage split from the ancestor). If a shared derived character, or synapomorphy, is found in two or more related organisms, it is inferred to have been present in their common ancestor, irrespective of whether or not there is a fossil record for that ancestor. Rather than trying to find the actual fossil corresponding to the “missing link” between lobe-fins and tetrapods, paleontologists instead look for fossils with characters or features important for an adaptive transition from life in an aquatic environment to life on land and that are shared as the result of common ancestry. [16]

Once again, one needs to emphasise that evolution is not a ladder, but a tree. Furthermore, evolutionary biologists do not look for missing links, but fossils with transitional features. Collyer’s use of the term “missing link” demonstrates that he is uninformed about the subject he criticises, making his criticisms invalid.
Fossils with transitional features (hereafter transitional fossils for brevity) will have characteristics of two distinct groups, and will give insight into what the common ancestor of these groups would resemble prior to the divergence of the two lines. For example, tetrapods are believed to have evolved from fish, so the common ancestor of tetrapods and fish will have fish-like and tetrapod-like features. Transitions at the species level generally are not abundant, but they are numerous at the higher taxonomic levels.
Jennifer Clack, who is a vertebrate palaeontologist with specific expertise in the evolution of tetrapods from fish-like ancestors, recently commented on the revolution in this field of palaeontology, with the discovery of new transitional fossils that have revolutionised our understanding of this area of evolution, as well as new interpretations of previously discovered fossils.
The idea that, once upon a time, creatures with fins left the water and crawled up onto the land is one that has pretty wide currency among the general public. It is recognized in many a cartoon, as well as being referred to in some form in a wide variety of media, and is often featured in books on prehistoric animals. However, the details are often only vaguely understood and often originate in ideas about the subject that were put forward in the early years of the twentieth century, based on very little in the way of hard facts. It was thus possible to misrepresent or lampoon scientific views of the subject, since speculation seemed to be in inverse proportion to the amount of data. In recent years, especially in the last 5 or 10 years, information and ideas about “the fish–tetrapod transition” have expanded and changed enormously, so that we can now refer to a wealth of fossil and other evidence to generate plausible and testable hypotheses. [105]
Clack’s opening remarks should remind special creationists to keep up to date with contemporary evolutionary biology, if only to avoid criticising evolutionary scenarios that are no longer held or advocated by mainstream biologists.
Why we should not expect the fossil record to have innumerable fine transitions
Collyer's claim that evolution requires many millions of missing links of a slow evolutionary process" immediately brings to mind the comment by Darwin, frequently quote mined and misunderstood by evolution denialists, about the absence of "innumerable transitional forms". What Collyer and other evolution denialists forget is that the fossil record reflects what one would expect under a model of allopatric speciation. As Douglas Theobald points out:
- There are two common uses of "gradualism," one of which is more traditional, the other of which is equivalent to Eldredge and Gould's "phyletic gradualism."
- Darwin was not a "phyletic gradualist," contrary to the claims of Eldredge and Gould.
- PE [punctuated equilibrium] is not anti-Darwinian; in fact, the scientific basis and conclusions of PE originated with Charles Darwin.
- PE does not require any unique explanatory mechanism (e.g. macromutation or saltation).
- Eldredge and Gould's PE is founded on positive evidence, and does not "explain away" negative evidence (e.g. a purported lack of transitional fossils). [18]
Theobald quotes palaeontologists Niles Eldredge and Stephen Jay Gould, the originators of Punctuated Equilibrium:
"In summary, we contrast the tenets and predictions of allopatric speciation with the corresponding statements of phyletic gradualism previously given:
(1) New species arise by the splitting of lineages.
(2) New species develop rapidly.
(3) A small sub-population of the ancestral form gives rise to the new species.
(4) The new species originates in a very small part of the ancestral species' geographic extent - in an isolated area at the periphery of the range.
These four statements again entail two important consequences:
(1) In any local section containing ancestral species, the fossil record for the descendant's origin should consist of a sharp morphological break between the two forms. .... we will rarely discover the actual event in the fossil record.
(2) Many breaks in the fossil record are real; they express the way in which evolution occurs, not the fragments of an imperfect record." (Eldredge and Gould 1972)
Given this, one would fully expect to see in the fossil record a relative absence of species-level transitions, but numerous transitions at higher taxonomic levels, exactly as Gould said.
What is fascinating is that Darwin himself made points quite similar to this in the Origin of Species. Theobald continues:
These tenets and conclusions of their "new" theory of PE are quite interesting when compared to these quotes from Origin of Species:
PE tenet #1 and #2"If, however, the modified offspring of a species get into some distinct country, or become quickly adapted to some quite new station, in which child and parent do not come into competition, both may continue to exist." (Darwin 1872, Ch. 4, "Natural Selection," p. 155)
This quote shows that Darwin understood tenet 1 above in the description of allopatric speciation. It also indicates that Darwin realized that species could develop rapidly (tenet 2 of PE), as he says they may "become quickly adapted." Also,
PE tenet #1,#2 #3, #4, and PE consequences #1 and #2
"... the periods during which species have undergone modification, though long as measured by years, have probably been short in comparison with the periods during which they retained the same form. It is the dominant and widely ranging species which vary most frequently and vary most, and varieties are often at first local—both causes rendering the discovery of intermediate links in any one formation less likely. Local varieties will not spread into other and distant regions until they are considerably modified and improved; and when they have spread, and are discovered in a geological formation, they appear as if suddenly created there, and will be simply classed as new species." (Darwin 1872, Ch. 15, "Recapitulation and Conclusion", p. 619)
This quote is remarkable in that it nearly states every tenet and prediction that Eldredge and Gould listed for their paleontological treatment of allopatric speciation. Darwin agreed with the other tenets of allopatric speciation, including tenets 3 and 4 - "varieties are often at first local" and "will not spread ... until they are considerably modified." Furthermore, it is obvious that Darwin agreed with the two predictions of the allopatric speciation model concerning what will be found in the paleontological record.
Here I present quotes from The Origin of Species , Chapter 10, "On the imperfection of the geological record," that basically sum up the conclusions of PE:
"When we see a species first appearing in the middle of any formation, it would be rash in the extreme to infer that it had not elsewhere previously existed. So again, when we find a species disappearing before the last layers have been deposited, it would be equally rash to suppose that it then became extinct. We forget how small the area of Europe is compared with the rest of the world ... when we see a species first appearing in any formation, the probability is that it only then first immigrated into that area." (p. 423)
"... varieties are generally at first local; and that such local varieties do not spread widely and supplant their parent-form until they have been modified and perfected in some considerable degree. According to this view, the chance of discovering in a formation in any one country all the early stages of transition between any two forms is small, for the successive changes are supposed to have been local or confined to some one spot." (pp. 427-428)
"... it might require a long succession of ages to adapt an organism to some new and peculiar line of life, for instance, to fly through the air; and consequently that the transitional forms would often long remain confined to some one region; but that, when this adaptation had once been effected, and a few species had thus acquired a great advantage over other organisms, a comparatively short time would be necessary to produce many divergent forms, which would spread rapidly and widely throughout the world." (p. 433)
It is obvious from all of these quotes that Darwin did not think the "gaps" between fossil species were only due to geological processes, but that they are a direct consequence of natural speciation processes. Phyletic gradualism is a strawman when attributed to Darwin, and this is one of the reasons why so many evolutionary biologists reacted strongly to the initial presentation of the hypothesis of PE. Furthermore, it is erroneous even to claim PE as an original concept, since all of the tenets of allopatric speciation and the conclusions labelled as PE were stated by Charles Darwin over 100 years before Eldredge and Gould proposed their "novel" hypothesis. [19]
Therefore, it should be clear that while evolution will produce a series of gradual transition forms, the fossil record will not likewise consist of an innumerable sequence of fine-graded transitions due to the mechanisms of speciation as outlined, coupled with the vagaries of the fossil record. Collyer once again shows that he simply did not understand the subject.
References
1. Gould S Evolution as Fact and Theory, Hen's Teeth and Horse's Toes: Further Reflections in Natural History, New York: W. W. Norton & Company, 1994, p. 260
2. Prothero D Evolution: What the Fossils Say and Why it Matters (2007: Columbia University Press)
3. Daeschler E.B., Shubin N.H, Jenkins F. “A Devonian tetrapod-like fish and the evolution of the tetrapod body plan.” Nature (2006) 440:757-634
4. Shubin NH, Daeschler EB, Jenkins FA. "The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb" Nature (2006) 440:764-71.
5. Downs JP, Drescher EB, Jenkins FA, Shubin NH "The cranial endoskeleton of Tiktaalik roseae" Nature (2008) 455:925-929
6. Shubin, NH, Daeschler EB, Jenkins FA "Pelvic girdle and fin of Tiktaalik roseae" Proc. Natl. Acad. Sci. USA (2014) 111:894-899
7. Thewissen JGM et al “From Land to Water: the Origin of Whales, Dolphins, and Porpoises” Evo Edu Outreach (2009) 2:272–288
8. Prothero DR "Evolutionary Transitions in the Fossil Record of Terrestrial Hoofed Mammals" Evo Edu Outreach (2009) 2:289–302
9. Smith FH, Cartmill M The Human Lineage (2009: John Wiley)
10. Chiappe LM "Downsized Dinosaurs: The Evolutionary Transition to Modern Birds" Evo Edu Outreach (2009) 2:248-256
11. Kemp TS The Origin and Evolution of Mammals (2005: Oxford University Press)
12. George TN "Fossils in Evolutionary Perspective," Science Progress (1960) 48:1-3
14. Romer AS “Cynodont Reptile with Incipient Mammalian Jaw Articulation” Science (1969) 166:881-882
15. Mead L.S. “Transforming Our Thinking About Transitional Forms” Evo Edu Outreach 2009 2:310-314
16. ibid, p 311
18. Theobald D "All You Need to Know About Punctuated Equilibrium (Almost)"
19. ibid


{kind=link}